高渗透压甘油蛋白激酶基因RkHog1及其重组表达载体的制作方法
本发明属于生物技术领域和遗传工程领域,涉及一种高渗透压甘油蛋白激酶基因rkhog1及其重组表达载体。
背景技术:
生物在长期的进化过程中形成了精确而敏感的适应机制,对外界环境刺激作出应答反应。有丝分裂原活化蛋白激酶(mitogen-activatedproteinkinase,mapk)是一类普遍存在于真核生物且在进化上保守的蛋白激酶,主要参与传导胞外信号,调节细胞的多种生物学过程。mapk通常由mapkk激活,而mapkk又受到mapkkk的活化。高渗透压甘油促分裂原激酶信号转导途径hog-mapk(highosmohrityglycerolmitogen-activatedproteinkinasesignalingtransductionpathway)是酿酒酵母对外界高渗透压胁迫环境应答的重要途径之一。该途径通过促进甘油积累、阻遏细胞生长及其他生理条件调节来抵抗渗透胁迫刺激。几乎所有真核生物的hog-mapk途径作用均是高渗透压环境中调控胞内生理生化反应以快速合成和/或积累胞内甘油提高胞内渗透压。
hog信号途径首先发现于酿酒酵母,该信号途径受到外界高渗条件的诱导,激活甘油合成相关酶基因(如gpd1和hor1)的转录,使胞内积累高浓度的甘油,维持细胞较高的膨压。近年来的研究表明,该信号途径广泛存在于其它丝状真菌,与真菌胞内的膨压调节密切相关。但在不同的真菌中,hog-mapk信号途径调节胞内小分子量多元醇的种类不同。如人类病原真菌(candidaalbicans)在高渗环境中,hog-mapk主要通过调控甘油的积累提高胞内膨压,抵御不良环境;植物病原真菌(magnaporthegrisea)中,当hog-mapk编码基因osm1被破坏后,菌体在高渗条件下的形态表现异常,胞内阿拉伯糖醇的积累显著降低,而甘油积累却没有明显变化。研究还发现,蛋白激酶hoglp与病原真菌的致病性有关。烟曲霉(a.fumigatus)osml与酵母hogl基因同源,osml基因突变体在高渗条件下毒力下降,菌丝形态改变;白念珠(c.albicans)hogl突变体菌丝形成受到抑制,突变株对小鼠的致死率显著下降。
近年来,hog-mapk途径的重要蛋白激酶hoglp的编码基因已在多种丝状真菌中被分离
和研究。红冬孢酵母(rhodosporidiumkratochvilovae)作为优良的微生物油脂生产菌,在营养限制条件下能过量积累胞内油脂;并生产胞外多糖、β-类胡萝卜素、工业用酶等,在生物制药产业和环境治理方面中有广泛的应用前景。同时,红冬孢酵母具有生长周期短、利用碳源广泛、油脂积累量多等优点,还能在高渗透压环境中生长,因此,该菌具有适应不同环境的调节机制。然而,有关hog信号途径与其环境适应性、生长发育、致病性等关系的研究较少。因此,克隆获得高渗透压甘油蛋白激酶基因rkhog1并进行功能研究,分析hog信号途径在该菌中的作用,明确hog信号途径的作用机理及其生长适应性的关系是为今后更好地利用及改造相关生物基因表达调控这一重要生物资源提供十分重要的手段,为利用基因工程技术改善微生物及其他高等真核生物的抗逆性改良提供了更加丰富的优良基因材料。
技术实现要素:
本发明的目的是提供一种高渗透压甘油蛋白激酶基因rkhog1,这是从红冬孢酵母(rhodosporidiumkratochvilovae)ym25235中分离得到,该基因核苷酸序列如seqidno:1所示或该核苷酸序列的片段,或与seqidno:1互补的核苷酸序列,该基因序列长为1080bp(碱基),该基因编码的氨基酸序列如seqidno:2所示的多肽或其片段。
本发明另一目的是提供一种含有高渗透压甘油蛋白激酶基因rkhog1的重组表达载体,是将seqidno:1所示基因直接与不同表达载体(质粒、病毒或运载体)连接所构建的重组载体。
本发明另一目的是提供一种含有高渗透压甘油蛋白激酶基因rkhog1或上述重组表达载体的宿主细胞酿酒酵母by4742。
具体涉及从红冬孢酵母(rhodosporidiumkratochvilovae)ym25235中克隆的高渗透压甘油蛋白激酶rkhog1基因及将该基因直接与不同载体连接,转入细菌、酵母或其他真菌中,并对其表达特性进行研究,为阐明hog途径在红冬孢酵母ym25235中的功能奠定了基础。
本发明从红冬孢酵母(rhodosporidiumkratochvilovae)ym25235的染色体基因组中分离得到hog途径的核心分子hog1p的基因rkhog1。该基因全长1080bp,研究表明基因rkhog1功能与酿酒酵母hog1完全相同,且性能更佳。该基因的获得扩大了微生物抗渗透压胁迫研究的基因资源,也为红冬孢酵母hog信号途径的分子机理奠定基础,为今后更好地利用及改造相关生物基因表达调控这一重要生物资源提供十分重要的手段。该基因是耐高渗透压调控的功能基因,若将耐高渗的功能基因hog1导入植物,就可以进行耐盐新品种的选育,又如利用hog1调控产甘油关键酶基因gpd高效启动子,可用于工业上产油菌株的改造;从而为培养耐高渗透压和高盐环境的生物新品种提供了新的材料和新的思路。
附图说明
图1为本发明的红冬孢酵母ym25235rkhog1基因pcr扩增图;
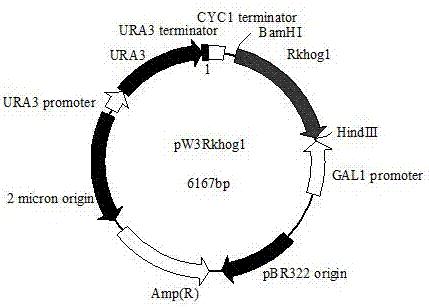
图2为利用本发明的红冬孢酵母ym25235rkhog1基因构建的酿酒酵母表达载体pw3rkhog1结构示意图;
图3为本发明所构建的重组表达质粒pw3rkhog1的酶切分析图;其中:1为dnamaker;2为空质粒pwxy3.0双酶切对照;3为重组质粒pw3rkhog1双酶切;4为rkhog1双酶切对照;5为dnamaker;
图4为重组菌株在不同nacl浓度的固体培养基上的菌落形态;a图为0.5mnacl浓度,b图为1.0mnacl浓度;图中从左到右分别为重组质粒pw3rkhog1转化的酿酒酵母hog1缺失菌株、重组质粒pw3schog1转化的酿酒酵母hog1缺失菌株、空质粒pwxy3.0转化的酿酒酵母hog1缺失菌株、空质粒pwxy3.0转化的酿酒酵母by4742野生菌株;
图5为重组菌株在不同山梨醇浓度的固体培养基上的菌落形态;a图为1.2m浓度,b图为1.8浓度;图中从左到右分别为重组质粒pw3rkhog1转化的酿酒酵母hog1缺失菌株、重组质粒pw3schog1转化的酿酒酵母hog1缺失菌株、空质粒pwxy3.0转化的酿酒酵母hog1缺失菌株、空质粒pwxy3.0转化的酿酒酵母by4742野生菌株。
具体实施方式
下面结合附图和实施例对本发明作进一步详细说明,但本发明保护范围不局限于所述内容,实施例中使用的试剂和方法,如无特殊说明,均采用常规试剂和使用常规方法。
实施例1:红冬孢酵母(rhodosporidiumkratochvilovae)ym25235rkhog1基因克隆
采用omega试剂盒e.z.n.afungalrnakit提取红冬孢酵母ym25235总rna,反转录试剂盒takarafirststrandcdnasynthesiskit合成cdna。根据红冬孢酵母ym25235的转录组序列设计特异性引物(引物1和引物2)进行pcr扩增。反应所用引物、组份和扩增条件如下:
引物1:rkhog1-f:5’-cgcaagcttatggccgacttcgtgaag-3’(seqidno:3)
引物2:rkhog1-r:5’-taatggatccttactgctgcggcgcg-3’(seqidno:4)
(aagctt为hindⅲ酶切位点,ggatcc为bamhⅰ酶切位点);
pcr扩增体系如下(50μl):
pcr扩增条件:95℃预变性5min,再用95℃变性30s,63℃退火15s,72℃延伸1min进行30个循环,最后72℃彻底延伸10min。反应完后取产物2μl,然后在浓度为1%的琼脂糖凝胶中,进行电泳分析,结果如图1所示。经凝胶成像系统成像确认片段大小正确后,用百泰克生物技术有限公司多动能dna纯化回收试剂盒回收目的片段,然后将pcr扩增得到的目的基因连接到pmd-18t上,连接产物转化至大肠杆菌dh5α感受态细胞中,用含有氨苄青霉素(amp+)的lb固体平板进行筛选,挑取平板上的转化子进行菌落pcr筛选阳性克隆,然后送去上海生工测序。测序结果显示,获得一段1080bp长的序列,命名为rkhog1,序列组成如seqidno:1所示的核苷酸序列。
实施例2:重组表达质粒pw3rkhog1的构建
采用实施例1中cdna为模板进行pcr扩增,反应所有引物组合、反应组分和扩增条件如下:
引物1:rkhog1-f:5’-cgcaagcttatggccgacttcgtgaag-3’(seqidno:3)
引物2:rkhog1-r:5’-taatggatccttactgctgcggcgcg-3’(seqidno:4)
(aagctt为hindⅲ酶切位点,ggatcc为bamhⅰ酶切位点);
pcr扩增体系如下(50μl):
pcr扩增条件:95℃预变性5min,再用95℃变性30s,63℃退火15s,72℃延伸1min进行30个循环,最后72℃彻底延伸10min。
将上述pcr获得的基因片段和表达载体pwxy3.0用hindⅲ和bamhⅰ核酸内切酶进行双酶切,用百泰克生物技术有限公司多动能dna纯化回收试剂盒回收酶切片段,并用t4连接酶在16℃连接过夜,连接产物转化大肠杆菌dh5α,用含有氨苄青霉素(amp+)的lb固体平板进行筛选,挑取平板上的转化子进行菌落pcr,筛选阳性克隆,同时进一步对转化子进行双酶切验证分析。结果显示,如图3第3泳道所示,用hindⅲ和bamhⅰ双酶切,重组质粒产生两条带,小分子条带与泳道4中该基因的pcr产物大小一致,大分子条带
与泳道2中用相同酶切质粒pwxy3.0后产生的条带大小一致,这表明所构建的重组质粒正确,进一步测序分析也证明这一点。将重组表达质粒命名为pw3rkhog1,该质粒图谱如图2所示。
实施例3:红冬孢酵母(rhodosporidiumkratochvilovae)ym25235rkhog1基因在提高酿酒酵母hog1缺失菌株中耐高渗的应用
1、重组质粒pw3rkhog1转化酿酒酵母hog1缺失菌株by4742
采用鲑鱼精dna介导的醋酸锂化学转化法将重组质粒pw3rkhog1转化至酿酒酵母hog1缺失菌株中,以尿嘧啶营养缺陷型(含2%葡萄糖)的培养基筛选转化子。菌落pcr验证转化子。
2、高渗条件下重组菌株的生长。
挑取空质粒转化的酿酒酵母by4742野生菌株、空质粒转化的酿酒酵母hog1缺失菌株、重组质粒pw3schog1转化的酿酒酵母hog1缺失菌株和重组质粒pw3rkhog1转化的酿酒酵母hog1缺失菌株单克隆于5ml的尿嘧啶营养缺陷型(含2%葡萄糖)液体培养基中,于30℃,150rpm培养24h;分别将上步酵母菌液于4000rpm下离心5min收集菌体,并用尿嘧啶营养缺陷型液体培养基洗涤3次,于4000rpm下离心5min收集菌体,于尿嘧啶营养缺陷型(含2%半乳糖+2%棉籽糖)液体培养基,并确保其初始od600值在0.3-0.6之间,在30℃,80rpm下诱导12h。分别将诱导后的菌液的od600值调整为0.8,并同时用血球计数板计数(四个菌株之间的菌数尽量调整一致,以避免较大的误差)。将上步od600值调整一致的菌液分别十倍稀释至10-1、10-2、10-3、10-4、10-5分别取2µl点板于尿嘧啶营养缺陷型(含2%葡萄糖)0.5mnacl、1.0mnacl、1.2m山梨醇、1.8m山梨醇平板上,28℃倒置培养48h。观察四个菌株菌落的差别(见图4和5)。(从左到右为重组质粒pw3rkhog1转化的酿酒酵母hog1缺失菌株、重组质粒pw3schog1转化的酿酒酵母hog1缺失菌株、空质粒pwxy3.0转化的酿酒酵母hog1缺失菌株、空质粒pwxy3.0转化的酿酒酵母by4742野生菌株)
通过rkhog1基因在酿酒酵母hog1缺失菌株by4742中互补表达可以看出,重组菌株by4742-rkhog1相较于对照原始菌株by4742耐高渗能力显著提高。
序列表
<110>昆明理工大学
<120>高渗透压甘油蛋白激酶基因rkhog1及其重组表达载体
<160>4
<170>patentinversion3.3
<210>1
<211>1080
<212>dna
<213>rhodosporidiumkratochvilovae
<400>1
atggccgacttcgtgaagctcagcatctttggaaccgtcttcgaggtcaccacccggtat60
gtcgacctccagccggtcggcatgggcgcgttcggactcgtctgctcggccaaggatgag120
ctgacaggccagatggtcgcgatcaagaagatcatgaagccgttcagcacccccgtcctc180
gcaaagcggacgtaccgcgagctcaagctcttgaagcacatccagcacgagaacatcatc240
tcgctgtccgacatcttcatctcccccctcgaggacatctactttgtcaccgagctgctc300
ggtaccgacctccaccgcctgttgacgagccggccgctcgagaagcagttcatccagtac360
ttcctctaccagattctccgtggcttgaagtacgtgcactcggcaggcgttgtccaccgc420
gacctcaagccctccaacatcctcgtcaacgagaactgcgacctcaagatctgcgacttt480
ggcctcgcgcgcctcgcagacccgcagatgaccggctacgtctcgacgcggtactaccgc540
gcgcctgagatcatgctcacgtggcagaagtacgacgttgccgtcgacatctggtccgcg600
gggtgcatcttcgccgagatgctggagggccgcccgctcttcccgggcaaggaccacgtc660
aaccagttctcgatcatcaccgagctgctggggacgccgcctgacgaggtcatccagacc720
atctgctcggagaacacccttcgcttcgtccagtcgctcccgaaacgcgagcgcgtcccg780
ttcgcgcagaagttccgcggcgcggacccgcaagcgctcgacctcctcgagaagatgctc840
gtgttcgacccgcgcaagcgcatcgacgcgacgcaggcgctcgcgcacgagtacctcgca900
ccgtaccacgacccgacggacgagcccgtcgcgcccgaggcgtttgactggagcttcaac960
gacgcggacctgccggtggacacgtggaaggtgatgatgtactcggagatcctcgacttc1020
caccacatcgaggacgggaccgagtcgggcccggtcgcgggcgccgcgccgcagcagtaa1080
<210>2
<211>360
<212>prt
<213>rhodosporidiumkratochvilovae
<400>2
metalaaspphevallysleuserilepheglythrvalphegluvalthrthrargtyr
11020
valaspleuglnprovalglymetglyalapheglyleuvalcysseralalysaspglu
3040
leuthrglyglnmetvalalailelyslysilemetlyspropheserthrprovalleu
5060
alalysargthrtyrarggluleulysleuleulyshisileglnhisgluasnileile
7080
serleuseraspilepheileserproleugluaspiletyrphevalthrgluleuleu
90100
glythraspleuhisargleuleuthrserargproleuglulysglnpheileglntyr
110120
pheleutyrglnileleuargglyleulystyrvalhisseralaglyvalvalhisarg
130140
aspleulysproserasnileleuvalasngluasncysaspleulysilecysaspphe
150160
glyleualaargleualaaspproglnmetthrglytyrvalserthrargtyrtyrarg
170180
alaprogluilemetleuthrtrpglnlystyraspvalalavalaspiletrpserala
190200
glycysilephealaglumetleugluglyargproleupheproglylysasphisval
210220
asnglnpheserileilethrgluleuleuglythrproproaspgluvalileglnthr
230240
ilecyssergluasnthrleuargphevalglnserleuprolysarggluargvalpro
250260
phealaglnlyspheargglyalaaspproglnalaleuaspleuleuglulysmetleu
270280
valpheaspproarglysargileaspalathrglnalaleualahisglutyrleuala
290300
protyrhisaspprothraspgluprovalalaproglualapheasptrpserpheasn
310320
aspalaaspleuprovalaspthrtrplysvalmetmettyrsergluileleuaspphe
330340
hishisilegluaspglythrgluserglyprovalalaglyalaalaproglngln***
350
<210>3
<211>27
<212>dna
<213>人工序列
<400>3
cgcaagcttatggccgacttcgtgaag27
<210>4
<211>26
<212>dna
<213>人工序列
<400>4
taatggatccttactgctgcggcgcg26
- 还没有人留言评论。精彩留言会获得点赞!