一种重组腺相关病毒的制备方法与流程
本发明属于基因治疗领域,更具体地,涉及一种重组腺相关病毒的制备方法。
背景技术:
重组腺相关病毒(raav)具有安全性高、免疫原性低、宿主范围广、能介导外源基因在动物体内长期稳定表达等特点,是基因治疗领域最具应用前景的载体之一。raav在神经科学研究以及疾病的基因治疗等领域具有重要作用和巨大需求。有研究数据表明,一次大型灵长类动物试验或临床基因治疗试验就需要约1015vg(vg,virusgenomes)的raav,是一般体外细胞试验或小鼠试验用量的成百上千倍(humgenether.2002nov1;13(16):1935-43.)。
杆状病毒(baculovirus)属于杆状病毒科,专一性寄生于节肢动物体内,多见于昆虫纲的鳞翅目昆虫,可感染600多种昆虫。杆状病毒是有囊膜的双链环状dna病毒,其基因组大小在90kb-230kb之间,编码上百种蛋白。1983年,smith首次利用苜蓿银纹夜蛾核型多角体病毒(autographacalifornicamulticapsidnuclearpolyhedrosisvirus,acmnpv)在草地贪夜蛾细胞系sf9中成功表达了人β-干扰素,首创了杆状病毒表达系统(molcellbiol,1983,3:2156-2165)。1985年,maeda用家蚕核型多角体病毒(bombyxmorinuclearpolyhydrosisvirus,bmnpv)作为表达载体,在家蚕体内表达了人α-干扰素(nature,1985,315:592-594)。此后,杆状病毒表达系统不断完善和发展,已经成为应用非常广泛的一种真核表达系统。昆虫是一类与腺相关病毒(adeno-associatedvirus,aav)同属于微小病毒科(parvoviridae)的浓核病毒 亚科(densovirinae)病毒的自然宿主。2002年,urabe等人证实了感染杆状病毒的sf9昆虫细胞可以支持aav的复制,利用分别携带有aav的rep基因、cap基因以及itr核心表达元件的三种重组杆状病毒共同感染sf9细胞制备出了raav,成功建立了三杆状病毒系统(threebacsystem),(humgenether.2002nov1;13(16):1935-43.)。在此基础上,研究者相继开发出了更适合大规模制备raav的两杆状病毒系统(twobacsystem),(molther.2008may;16(5):924-30,molther.2009nov;17(11):1888-96.)以及依赖感染诱导表达rep基因和cap基因的包装细胞系的一杆状病毒系统(onebacsystem),(procnatlacadsciusa.2009mar31;106(13):5059-64,humgenether.2014mar;25(3):212-22.)。
目前利用杆状病毒系统大规模制备raav的方法,都是依靠重组杆状病毒感染体外培养的特定的昆虫细胞系,如sf9、sf21细胞等。然而体外细胞系培养工艺复杂,设备条件要求苛刻、生产成本高昂。
技术实现要素:
针对现有技术的以上缺陷或改进需求,本发明提供了一种重组腺相关病毒的制备方法,其目的在于利用一种重组杆状病毒感染昆虫幼虫大规模生产重组腺相关病毒,由此解决现有的重组腺相关病毒生产方法工艺复杂、条件要求苛刻、生产成本高昂的技术问题。
为实现上述目的,按照本发明的一个方面,提供了一种重组腺相关病毒的制备方法,其特征在于,包括以下步骤:
(1)将目标基因构建在重组型杆状病毒上,所述重组杆状病毒其基因组整合了制备重组腺相关病毒所必需的腺相关病毒的rep基因、cap基因以及携带有目标基因的itr核心表达元件;
(2)将步骤(1)中制得的重组杆状病毒感染宿主昆虫幼虫,使得所述宿主昆虫幼虫体内产生大量重组腺相关病毒;
(3)将步骤(2)中获得的宿主昆虫幼虫裂解,提取重组型腺相关病 毒并进行纯化。
优选地,所述制备方法,其步骤(2)具体包括以下步骤:
(2-1-1)将孵化后的宿主昆虫幼虫饲养至幼虫长到4-5龄;
(2-1-2)将步骤(1)中制得的重组杆状病毒,对(2-1-1)中获得的宿主昆虫幼虫进行皮下注射接毒,滴度在1×10e+7vg/ml至1×10e+9vg/ml,接毒体积在10μl-20μl;
(2-1-3)将步骤(2-1-2)中注射了重组杆状病毒的宿主昆虫幼虫饲养至幼虫出现食欲减弱、行动迟缓、体节肿胀、和/或体色变淡的病变症状时,结束饲养收获虫体。
优选地,所述制备方法,其步骤(2-1-3)在所述宿主昆虫幼虫感染相应所述重组杆状病毒后的3至7天期间出现典型病变症状时,虫体死亡之前结束饲养。
优选地,所述制备方法,其所述重组型杆状病毒为基于苜蓿银纹夜蛾核型多角体病毒(acmnpv)或家蚕核型多角体病毒(bmnpv)。
优选地,所述制备方法,其所述宿主昆虫幼虫为甜菜夜蛾(beetarmyworm,spodopteraexigua)幼虫或家蚕(silkworm,bombyxmori)幼虫。
优选地,所述制备方法,其所述步骤(3)将所述宿主昆虫幼虫裂解,其具体操作步骤如下:
将所述宿主昆虫幼虫捣碎研磨,加入pbs溶液匀浆,冻融后,制得裂解物。
优选地,所述制备方法,其步骤(3)所述提纯具体操作如下:
将所述宿主昆虫幼虫裂解后,将裂解物离心,取上清液,用氯仿抽提,获得初步纯化的重组腺相关病毒。
总体而言,通过本发明所构思的以上技术方案与现有技术相比,能够取得下列有益效果:
饲养昆虫幼虫相比体外培养昆虫细胞,具有设备简单容易推广,成本 低,产物生物活性高,是天然的生物反应器产量高等不可比拟的优势。本发明提供的方法,通过一种高度整合的重组杆状病毒感染昆虫幼虫,从而大量制备品质稳定的基因治疗用重组腺相关病毒载体。
附图说明
图1是本发明提供的重组腺相关病毒制备方法的流程示意图;
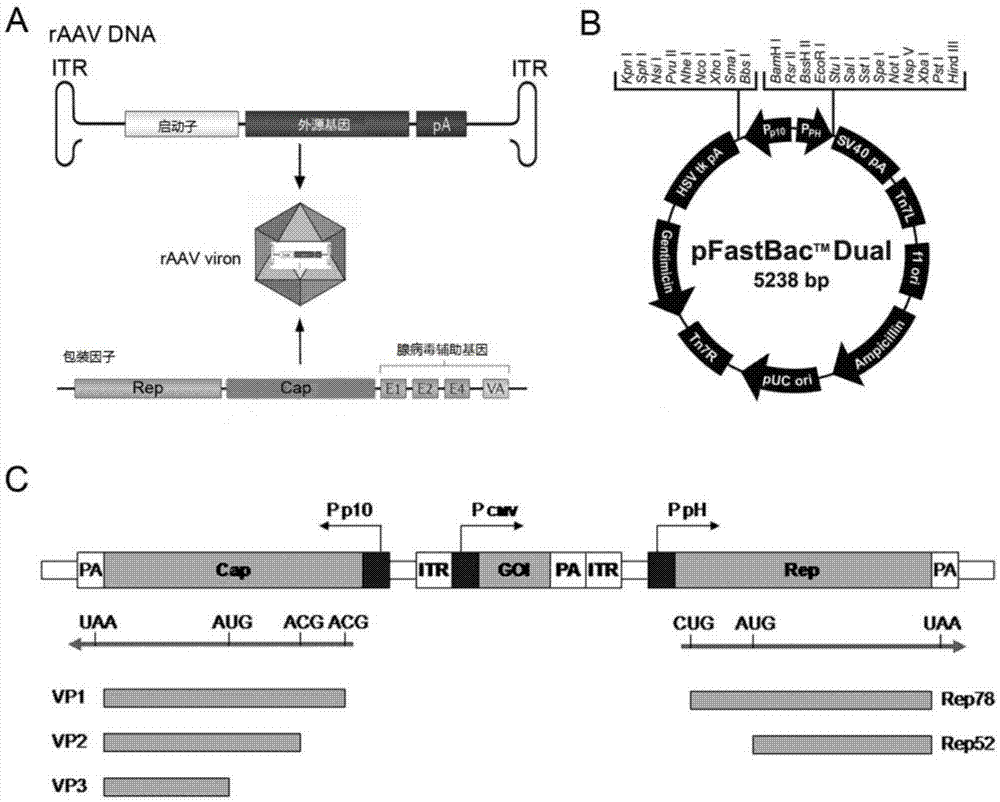
图2是raav的包装原理图(a),杆状病毒表达系统(bactobac)中穿梭质粒pfast.bac.dual(pfbd)的结构示意图(b)以及整合了制备raav所需组件的重组穿梭质粒pfbd/cap-(itr-gfp)-rep的结构示意图(c);
图3是实施例中利用重组杆状病毒感染宿主昆虫细胞或宿主昆虫幼虫制备raav的过程示意图;
图4是实施例1中制备重组杆状病毒感染宿主sf9细胞制备raav以及病毒活性的验证图;其中,图4a是重组杆状病毒bev/cap-(itr-gfp)-rep感染sf9细胞后的荧光显微成像图;图4b是实例1中制备的raav感染hek293和sf9细胞后的荧光显微成像图;
图5是实施例1中使用的正常甜菜夜蛾幼虫以及感染了重组杆状病毒后出现明显病变的虫体形态图。
图6是实施例1中利用重组杆状病毒感染宿主甜菜夜蛾幼虫制备raav以及病毒活性的验证图;其中,图6a是虫体裂解液上清感染hek293和sf9细胞后的荧光显微成像图;图6b是氯仿抽提纯化后的裂解液上清感染hek293细胞的荧光显微成像图;
图7是实施例2中制备重组杆状病毒感染宿主bmn细胞制备raav以及病毒活性的验证图;其中,图7a是重组杆状病毒bev/cap-(itr-gfp)-rep感染bmn细胞后的荧光显微成像图;图7b是实例2中制备的raav感染hek293和bmn细胞后的荧光显微成像图;
图8是实施例2中使用的正常家蚕幼虫以及感染了重组杆状病毒后出现明显病变的虫体形态图。
图9是实施例2中利用重组杆状病毒感染宿主家蚕幼虫制备raav以及病毒活性的验证图;其中,图9a是虫体裂解液上清感染hek293和bmn细胞后的荧光显微成像图;图9b是氯仿抽提纯化后的裂解液上清感染hek293细胞的荧光显微成像图。
具体实施方式
为了使本发明的目的、技术方案及优点更加清楚明白,以下结合附图及实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅仅用以解释本发明,并不用于限定本发明。此外,下面所描述的本发明各个实施方式中所涉及到的技术特征只要彼此之间未构成冲突就可以相互组合。
昆虫是杆状病毒的天然宿主。然而,目前利用杆状病毒系统大规模制备raav的方法,都是依靠感染体外培养的特定的昆虫细胞系。昆虫幼虫饲养相比体外培养昆虫细胞,具有设备简单容易推广,成本低,产物生物活性高,是天然生物反应器产量高等不可比拟的优势。迄今,尚没有利用重组杆状病毒感染昆虫幼虫制备raav的方法。原因是目前的杆状病毒系统制备raav方法存在难以克服的缺陷:现有的三杆状病毒系统或两杆状病毒系统,由于多种重组杆状病毒共感染单个昆虫细胞是随机事件效率低,感染昆虫幼虫相比体外培养的昆虫细胞将更加无法控制,raav的高效生产无法实现。对于现有的依赖感染诱导表达rep基因和cap基因的包装细胞系的一杆状病毒系统,虽然将raav的包装元件rep基因、cap基因构建到包装细胞系基因组中可行,然而构建类似的诱导表达rep基因、cap基因的转基因昆虫幼虫却难以实现。尚没有利用重组杆状病毒感染昆虫幼虫制备raav的应用。开发出不依赖包装细胞系的制备raav的真正的一杆状病毒系统(onebacsystem),是实现利用重组杆状病毒感染昆虫大规模制备raav的关键前提条件,如图3所示。
优选采用苜蓿银纹夜蛾核型多角体病毒或家蚕核型多角体病毒。
苜蓿银纹夜蛾核型多角体病毒(acmnpv)是一种能交叉感染30多种鳞翅目昆虫的杆状病毒,由于其感染宿主存在特异性,acmnpv不感染家蚕,而能够感染甜菜夜蛾。由于甜菜夜蛾野外分布广泛,容易人工饲养,是研究acmnpv感染昆虫虫体的常用宿主昆虫之一(plosone.2012;7(7):e42462),基于acmnpv的杆状病毒表达系统是开发最早,应用最广泛的一种杆状病毒表达系统(biotechnolj.2015may;10(5):702-14.)。
家蚕核型多角体病毒(bmnpv)感染宿主范围较小,其不感染甜菜夜蛾,而能够感染家蚕。家蚕是一种重要的经济昆虫,传统农业中种桑家蚕获得蚕丝并生产丝绸,在世界上有着上千年的悠久历史。基于bmnpv的家蚕杆状病毒表达系统也已经发展成为一种应用广泛的外源蛋白表达系统(applmicrobiolbiotechnol.2010jan;85(3):459-70)。
研发利用重组杆状病毒感染昆虫大规模制备raav的新方法,将为重要基因治疗病毒载体之一的raav生产提供崭新的平台,也将促进生物医药经济中的基因治疗产业和传统农业经济中的昆虫养殖业的融合发展。
本发明提供的重组腺相关病毒的制备方法,如图1所示,包括以下步骤:
(1)将目标基因构建在重组型杆状病毒上,所述重组型杆状病毒其基因组整合了腺相关病毒的rep基因、cap基因以及携带有目标基因的itr核心表达元件。
所述重组杆状病毒,优选基于acmnpvclonee2(基因组序列如:genbankaccessionno.km667940.1,参考journalofvirology,aug.1993,p.4566-4579)或acmnpvclonec6(基因组序列如:genbankaccessionno.nc_001623.1)或bmnpvisolatet3(基因组序列如:genbankaccessionno.l33180.1,参考virologicasinica,june2007,22(3):0218-225),整合了aav的rep基因、cap基因和携带有目标基因的itr核心表达元件,所述rep基因和cap基因优选依据核糖体泄露扫描原理进行了密码子优化。
所述itr核心表达元件,两端包括aav基因组中的一段末端反向重复序列(itr),中间为携带的目标基因(goi)片段,通过5’端连接核酸片段和3’端连接核酸片段与所述rep基因和cap基因的表达框相连。其中,rep基因和cap基因,皆可处于itr核心表达元件的上游或下游。
所述rep基因依据核糖体泄露扫描原理进行了密码子优化,通过ph启动子转录出一条mrna,实现rep72、rep52复制蛋白的功能性表达。其序列优选为如seqidno.1所示。
所述cap基因密码子优化同样利用核糖体泄露扫描原理,通过p10启动子转录出一条mrna,实现vp1、vp2、vp3三种衣壳蛋白以接近天然比例(1:1:10)的功能性表达。其序列优选为如seqidno.2所示。
itr核心表达元件,通过两端的连接核酸片段与rep基因、cap基因的表达框连接。itr为aav基因组中的一段末端反向重复序列,优选2型aav的itr序列,如seqidno.3所示。所述5’端或3’端连接核酸序列优选长度在80bp-140bp之间的连接核酸序列,如seqidno.4或seqidno.5所示。itr核心表达元件在实例中选用含cmv启动子、gfp基因、sv40ploya组件的gfp基因表达框,便于技术方案的验证。
整合到杆状病毒基因组上的rep基因、cap基因和itr核心表达元件的优选组合方案,如图2c所示。
本发明提供的重组杆状病毒(优选基于acmnpv或bmnpv),可按照如下方法制备:
a、采用基因人工合成的方法获得密码子优化的rep基因、cap基因;采用基因人工合成、pcr扩增和酶切连接的方法获得itr核心表达元件。
b、通过分子克隆的方法将步骤a中获得的rep基因、cap基因以及itr核心表达元件构建到pfast.bac.dual(pfbd)穿梭载体上(如图2b),根据bactobac系统操作方法,将重组杆状病毒基因组转染相应的宿主sf9细胞或bmn细胞,获得相应的重组杆状病毒。
raav载体dna一般是用外源目的基因表达元件替换aav的编码基因,仅保留了病毒复制和包装所需的itr序列。制备raav需要反式补偿rep基因、cap基因和辅助病毒功能因子,如图2a所示。
(2)将步骤(1)中制得的重组型杆状病毒感染宿主昆虫幼虫,使得所述宿主昆虫幼虫体内产生大量重组腺相关病毒。所述感染可通过皮下注射接毒方式进行,具体包括以下步骤:
(2-1-1)将孵化后的宿主昆虫幼虫,在适宜温度和湿度的洁净培养空间中,利用饲料饲养,直至幼虫长到4-5龄。
(2-1-2)将制得的重组杆状病毒,以适当病毒剂量,对4-5龄宿主昆虫幼虫进行皮下注射接毒。
(2-1-3)继续饲养感染了重组杆状病毒的宿主昆虫幼虫,使病毒在幼虫体内复制增值,直至病毒大量复制后幼虫出现食欲减弱、行动迟缓、体节肿胀、和/或体色变淡的典型病变症状时时,一般在注射后3-7天,结束饲养收获虫体。
(3)将步骤(2)中获得的昆虫幼虫裂解,提取重组型腺相关病毒并进行纯化。
将收集的虫体捣碎研磨,加入pbs溶液匀浆,然后反复冻融3次,5000rpm离心10min,取虫体裂解上清液。然后用氯仿抽提,获得初步纯化的raav。
以下为实施例:
实施例1.重组杆状病毒bev/cap-(itr-gfp)-rep感染甜菜夜蛾幼虫制备raav
(1)重组杆状病毒bev/cap-(itr-gfp)-rep的构建及其活性验证
为了将raav制备所需的三种主要组件:cap基因、rep基因以及含有itr的核心表达元件置于一个重组杆状病毒中。我们利用了杆状病毒表达系统bactobac中的pfast.bac.dual(pfbd)穿梭载体。实例中基于2型aav 的rep基因依据核糖体泄露扫描原理进行了密码子优化,rep基因置于ph启动子的调控之下,实现rep72、rep52复制蛋白的功能性表达。rep基因序列为如seqidno.1所示的序列。实例中基于2型aav的cap基因依据核糖体泄露扫描原理进行了密码子优化,cap基因置于p10启动子的调控之下,实现vp1、vp2、vp3三种衣壳蛋白以接近天然比例(1:1:10)的功能性表达。cap基因序列为如seqidno.2所示的序列。itr核心表达元件,实例中itr选用2型aav的itr核酸序列,即如seqidno.3所示的序列,itr的核心表达元件采用了绿色荧光蛋白(gfp)表达框,由cmv启动子控制gfp的表达,便于检测raav的活性。itr核心表达元件通过5’端连接核酸片段和3’端连接核酸片段与rep基因和cap基因的表达框相连。5’端或3’端连接核酸片段为如seqidno.4(linka)或seqidno.5(linkb)所示的序列。
在本实例中,重组杆状病毒中主要组件选取的组合方案如下;
cap-linka-(itr-gfp)-linkb-rep
利用常规分子克隆技术将itr核心表达元件(itr-gfp)通过连接核酸片段置于pfbd载体的p10和ph启动子之间,构建pfbd/cap-(itr-gfp)-rep重组穿梭质粒,如图2c所示。
参照bactobac系统操作说明,将该重组穿梭质粒转化含有acmnpv杆状病毒基因组的acdh10bac菌。通过tn7转座子元件介导的重组得到重组杆状病毒基因组(bacmid)。通过蓝白斑筛选和pcr鉴定获得含有重组bacmid的阳性菌。抽提纯化重组bacmid,将其转染贴壁培养的sf9细胞。昆虫细胞逐渐被转染后产生的重组杆状病毒完全感染并出现明显的细胞病变(cpe)现象。将细胞培养液3000rpm离心5min,上清中就含有产生的重组杆状病毒。
将上清液感染贴壁培养的sf9细胞后继续培养3天,可以看到对照组未感染病毒的sf9细胞状态正常,没有gfp的表达。而在实验组感染了bev 的sf9细胞出现了明显的cpe现象,有明显的gfp表达,如图4a。这表明该方案成功制备出了重组杆状病毒。将转染sf9细胞后制备的bev再感染培养的sf9细胞,感染3天后细胞出现明显cpe,将细胞培养液3000rpm离心5min,收上清即可获得大量bev。病毒的滴度用荧光定量pcr的方法进行测定,参考文献(procnatlacadsciusa,2009.106(13):p.5059-64)中的方法。
将实施例1中制备得到的bev以感染复数(moi=5)感染培养的sf9细胞,感染72小时后,分别收集培养上清和细胞。bev产生后,主要通过分泌释放到培养基上清中,在sf9细胞中也有部分未释放的bev。raav产生后,主要存在于细胞核中,由于昆虫细胞被感染后发生细胞病变(cpe),部分细胞会裂解,raav也会有部分释放到上清中。而因此上清和细胞中会同时存在bev和raav。
为了验证用重组杆状病毒感染sf9细胞制备出了具有活性的raav。在此通过病毒感染hek293细胞和sf9细胞的实验方法证实该系统制备出了raav,实验结果如图4b。具体实施过程和结果分析如下:将细胞沉淀反复冻融3次裂解后,5000rpm离心5min收集细胞裂解上清液。因为raav无包膜,经60℃、30分钟处理,活性不受影响;而重组杆状病毒(bev)有包膜,经60℃、30分钟处理,失去活性。含有病毒的上清液感染hek293细胞2天后,利用传统的三质粒转染hek293细胞方法制备的raav对照组,在处理前后都有gfp表达,表明raav在处理前后都有感染活性。bev感染sf9细胞后的培养基上清组,未处理的有较强gfp表达,处理后仅有少量gfp表达,表明bev可以感染hek293细胞,灭活处理后bev不感染,但上清中少量的raav仍能够感染。细胞沉淀裂解液上清实验组,未处理的有较强gfp表达,处理后gfp强度有降低但仍有较强荧光,表明灭活处理后bev不感染,但上清中大量的raav仍能够感染。用与感染hek293细胞组相同的样品感染sf9细胞2天后,结果显示:利用传统的 三质粒转染hek293细胞方法制备的raav对照组,没有gfp荧光,表明raav对sf9细胞没有感染性。bev感染sf9细胞后的培养基上清组,灭活后前有较强gfp表达,灭活后的没有gfp表达。细胞沉淀裂解液上清实验组,未处理的有较强gfp表达,处理后的没有gfp表达。
图4以上结果证明,重组杆状病毒bev/cap-(itr-gfp)-rep感染培养的sf9细胞能够制备出raav。
(2)利用重组杆状病毒(bev)感染甜菜夜蛾幼虫制备raav
将孵化后的甜菜夜蛾幼虫饲养至4-5龄,长度约2cm左右(虫重约0.15g),将虫体在冰上麻醉,然后用微量注射器对虫体进行皮下注射接毒,注射量约10μl(bev的滴度约为1×10e+9vg/ml)的重组杆状病毒,然后放入有盖的培养皿中,继续饲养3-4天,被感染的宿主幼虫明显出现食欲减弱,行动迟缓的病变现象。典型症状为体结肿胀,体色变淡,表皮薄脆。未感染的甜菜夜蛾幼虫状态正常(如图5)。
(3)裂解虫体,抽提raav并验证其活性
将收集的虫体捣碎研磨,加入pbs溶液匀浆,然后反复冻融3次,5000rpm离心10min,取虫体裂解上清液。bev感染甜菜夜蛾虫体后,bev增殖的同时,虫体细胞也将产生raav。虫体裂解上清液中会同时存在bev和raav。
为了验证用重组杆状病毒注射感染甜菜夜蛾幼虫制备出了具有活性的raav。通过病毒感染hek293细胞和sf9细胞的实验方法证实在此制备出了raav,实验结果如图6a。因为raav无包膜,经60℃、30分钟处理,活性不受影响;而重组杆状病毒(bev)有包膜,经60℃、30分钟处理,失去活性。具体实施过程和结果分析如下:
含有病毒的上清液感染hek293细胞2天后,注射感染bev的虫体裂解上清液实验组,未处理的有较强gfp表达,处理后gfp强度有降低但仍有较强荧光,表明灭活处理后bev不感染,但上清中大量的raav仍能 够感染。未感染的虫体裂解上清液对照组,在处理前后都没有gfp表达。
含有病毒的上清液感染sf9细胞2天后,注射感染bev的虫体裂解上清液实验组,未处理的有gfp表达,处理后没有gfp表达,表明bev在虫体中增殖,灭活后失去感染性,而raav不感染sf9细胞。未感染的虫体裂解上清液对照组,在处理前后都没有gfp表达。
将虫体裂解上清液,用氯仿抽提,进行初步纯化。因为raav是无囊膜的病毒,能够耐受氯仿处理,仍旧保持感染活性;而重组杆状病毒acmnpv是有囊膜的病毒,不耐受氯仿处理,失去活性。将氯仿处理初步纯化过的虫体裂解上清液感染hek293细胞,感染2天后,可以看到gfp的表达,如图6b所示。以上结果证明,重组杆状病毒bev/cap-(itr-gfp)-rep感染甜菜夜蛾幼虫,成功制备出了具有活性的raav。
实施例2.重组杆状病毒bev/cap-(itr-gfp)-rep感染家蚕幼虫制备raav
(1)重组杆状病毒bev/cap-(itr-gfp)-rep的构建及其活性验证
本实施例与实施例1关于构建重组杆状病毒过程中的其主要组件组合方案相似,如下:
cap-linka-(itr-gfp)-linkb-rep
利用常规分子克隆技术将itr核心表达元件(itr-gfp)通过连接核酸片段置于pfbd载体的p10和ph启动子之间,构建pfbd/cap-(itr-gfp)-rep重组穿梭质粒,如图2c所示。
参照bactobac系统操作说明,将该重组穿梭质粒转化含有bmnpv杆状病毒基因组的bmdh10bac菌。通过tn7转座子元件介导的重组得到重组杆状病毒基因组(bacmid)。通过蓝白斑筛选和pcr鉴定获得含有重组bacmid的阳性菌。抽提纯化重组bacmid,将其转染贴壁培养的bmn细胞。昆虫细胞逐渐被转染后产生的重组杆状病毒完全感染并出现明显的细胞病变(cpe)现象。将细胞培养液3000rpm离心5min,上清中就含有产生的 重组杆状病毒。
将上清液感染贴壁培养的bmn细胞。后继续培养3天,可以看到对照组未感染病毒的bmn细胞状态正常,没有gfp的表达。而在实验组感染了bev的bmn细胞出现了明显的cpe现象,有明显的gfp表达,如图7a。这表明该方案成功制备出了重组杆状病毒。将转染bmn细胞后制备的bev再感染培养的bmn细胞,感染3天后细胞出现明显cpe,将细胞培养液3000rpm离心5min,收上清即可获得大量bev。病毒的滴度用荧光定量pcr的方法进行测定,参考文献(procnatlacadsciusa,2009.106(13):p.5059-64)中的方法。
将实施例2中制备得到的bev以感染复数(moi=5)感染培养的bmn细胞,感染72小时后,分别收集培养上清和细胞。bev产生后,主要通过分泌释放到培养基上清中,在bmn细胞中也有部分未释放的bev。raav产生后,主要存在于细胞核中,由于昆虫细胞被感染后发生细胞病变(cpe),部分细胞会裂解,raav也会有部分释放到上清中。而因此上清和细胞中会同时存在bev和raav。
为了验证用重组杆状病毒感染bmn细胞制备出了具有活性的raav。在此通过病毒感染hek293细胞和bmn细胞的实验方法证实该系统制备出了raav,实验结果如图7b。具体实施过程和结果分析如下:将细胞沉淀反复冻融3次裂解后,5000rpm离心5min收集细胞裂解上清液。因为raav无包膜,经60℃、30分钟处理,活性不受影响;而重组杆状病毒(bev)有包膜,经60℃、30分钟处理,失去活性。含有病毒的上清液感染hek293细胞2天后,利用传统的三质粒转染hek293细胞方法制备的raav对照组,在处理前后都有gfp表达,表明raav在处理前后都有感染活性。bev感染bmn细胞后的培养基上清组,未处理的有较强gfp表达,处理后仅有少量gfp表达,表明bev可以感染hek293细胞,灭活处理后bev不感染,但上清中少量的raav仍能够感染。细胞沉淀裂解液上清实验组, 未处理的有较强gfp表达,处理后gfp强度有降低但仍有较强荧光,表明灭活处理后bev不感染,但上清中大量的raav仍能够感染。用与感染hek293细胞组相同的样品感染bmn细胞2天后,结果显示:利用传统的三质粒转染hek293细胞方法制备的raav对照组,没有gfp荧光,表明raav对bmn细胞没有感染性。bev感染bmn细胞后的培养基上清组,灭活后前有较强gfp表达,灭活后的没有gfp表达。细胞沉淀裂解液上清实验组,未处理的有较强gfp表达,处理后的没有gfp表达。
图7以上结果证明,重组杆状病毒bev/cap-(itr-gfp)-rep感染培养的bmn细胞能够制备出raav。
(2)利用重组杆状病毒(bev)感染家蚕幼虫制备raav
将孵化后的家蚕幼虫饲养至4-5龄,长度约4cm左右(虫重约1g),将虫体在冰上麻醉,然后用微量注射器对虫体进行皮下注射接毒,注射量约20μl(bev的滴度约为1×10e+7vg/ml)的重组杆状病毒,然后放入有盖的培养皿中,继续饲养6-7天,被感染的家蚕幼虫明显出现食欲减弱,行动迟缓的病变现象。典型症状为,体结肿胀,体色变淡,表皮薄脆。而未感染的家蚕幼虫状态正常,如图8所示。
(3)裂解虫体,抽提raav并验证其活性
将收集的虫体捣碎研磨,加入pbs溶液匀浆,然后反复冻融3次,5000rpm离心10min,取虫体裂解上清液。bev感染家蚕幼虫后,bev增殖的同时,虫体细胞也将产生raav。虫体裂解上清液中会同时存在bev和raav。
为了验证用重组杆状病毒注射感染家蚕幼虫制备出了具有活性的raav。通过病毒感染hek293细胞和bmn细胞的实验方法证实在此制备出了raav,实验结果如图9a。因为raav无包膜,经60℃、30分钟处理,活性不受影响;而重组杆状病毒(bev)有包膜,经60℃、30分钟处理,失去活性。具体实施过程和结果分析如下:
含有病毒的上清液感染hek293细胞2天后,注射感染bev的虫体裂解上清液实验组,未处理的有较强gfp表达,处理后gfp强度有降低但仍有较强荧光,表明灭活处理后bev不感染,但上清中大量的raav仍能够感染。未感染的虫体裂解上清液对照组,在处理前后都没有gfp表达。
含有病毒的上清液感染bmn细胞2天后,注射感染bev的虫体裂解上清液实验组,未处理的有gfp表达,处理后没有gfp表达,表明bev在虫体中增殖,灭活后失去感染性,而raav不感染bmn细胞。未感染的虫体裂解上清液对照组,在处理前后都没有gfp表达。
将虫体裂解上清液,用氯仿抽提,进行初步纯化。因为raav是无囊膜的病毒,能够耐受氯仿处理,仍旧保持感染活性;而重组杆状病毒bmnpv是有囊膜的病毒,不耐受氯仿处理,失去活性。将氯仿处理初步纯化过的虫体裂解上清液感染hek293细胞,感染2天后,可以看到gfp的表达,如9b所示。
以上结果证明,重组杆状病毒bev/cap-(itr-gfp)-rep感染家蚕幼虫,成功制备出了具有活性的raav。
本领域的技术人员容易理解,以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!