一种提高植物对抗有害昆虫能力的基因及其应用的制作方法
本发明属于分子生物学和植物学领域,更具体地,本发明涉及一种提高植物对抗有害昆虫能力的基因及其应用。
背景技术:
植物在生长发育的过程中遭受着各种各样的植食性昆虫的威胁,所以植物需要一套非常复杂的防御体系来抵抗这些昆虫的侵害。目前,植物如禾本科植物对昆虫(如飞虱科昆虫)的抗性分子机制的研究还处于初级阶段,发掘与昆虫抗性相关的基因,增强对昆虫防御分子机制的了解,将有助于田间对于昆虫的防治。
植物体内的代谢产物按其生物学功能可分为初生代谢产物(primarymetabolites)和次生代谢产物(secondarymetabolites)两大类。次生代谢物质的合成是受细胞内部相关基因调控的一系列复杂的生化反应过程,因此在植物细胞内必然存在着相关的胞内信号分子和相应的信号转导机制来感受并传递外界因子的刺激信号。抗氧化类黄酮类是普遍分布于植物中的一类植物次生代谢物。根据其分子结构及杂环吡喃环的构象可将其分为6类:黄酮、黄烷酮、异黄酮、黄酮醇、黄烷醇和花色素苷。其中异黄酮和花色素苷作为目前研究的热点,主要在于大量的研究证实,这类化合物具有抗氧化、清除自由基、保护心脑血管、抗肿瘤和抗炎症等多种药用价值。在植物体内,异黄酮产物能够增加植物在抗病反应中的抗性,是具有广谱抗菌活性的植物保护素;同时是豆科植物和固氮菌之间的信号分子。黄酮类化合物在植物体内常与糖类结合成苷类,即花色素苷,或以游离的形式存在于植物液泡中,使花、茎、叶、果等器官呈现各种颜色,可吸引昆虫传粉,使果实免受紫外光伤害;且花色素苷根据种类和数量的不同使种或品种表现不同颜色,对植物的生长发育、开花结果以及防御侵害具有重要作用。
技术实现要素:
本发明的目的在于提供一种提高植物对抗有害昆虫能力的基因及其应用。
在本发明的第一方面,提供一种调节植物对抗昆虫的能力或调节植物花青素含量的方法,所述方法包括:调节植物内黄烷酮3-羟化酶(f3h)的表达或活性。
在一个优选例中,所述的方法包括:上调植物中黄烷酮3-羟化酶的表达或活性,从而提高植物对抗昆虫的能力,或提高植物的花青素含量。
在另一优选例中,所述的上调植物中黄烷酮3-羟化酶的表达或活性包括:将黄烷酮3-羟化酶的过量表达分子转入植物细胞、组织、器官或种子中,从而上调植物中黄烷酮3-羟化酶的表达或活性。
在另一优选例中,所述的方法包括:下调植物中黄烷酮3-羟化酶的表达或活性,从而降低植物对抗昆虫的能力,或降低植物的花青素含量。
在另一优选例中,所述的下调植物中黄烷酮3-羟化酶的表达或活性包括:在植物的基因组中敲除或沉默黄烷酮3-羟化酶编码基因;或将下调黄烷酮3-羟化酶编码基因的转录或表达的下调剂转入植物中;较佳地,所述的下调黄烷酮3-羟化酶编码基因的转录或表达的下调剂是特异性干扰黄烷酮3-羟化酶基因表达的干扰分子;更佳地,所述的干扰分子是以黄烷酮3-羟化酶基因或其转录本为抑制或沉默靶标的dsrna,反义核酸,或shrna,或能表达或形成所述dsrna,反义核酸或shrna的构建物。
在另一优选例中,所述的特异性干扰黄烷酮3-羟化酶编码基因表达的干扰分子以黄烷酮3-羟化酶编码基因中第500-981位的核酸片段作为靶标。
在另一优选例中,所述的植物包括双子叶植物和单子叶植物;较佳地,所述的植物包括禾本科植物,十字花科植物,茄科植物或大戟科植物。
在另一优选例中,所述的十字花科植物包括:拟南芥;或者所述的禾本科植物包括:水稻、小麦、玉米;或者所述的茄科植物包括:马铃薯、西红柿;或者所述的大戟科植物包括:木薯。
在另一优选例中,所述的昆虫是飞虱科昆虫,同翅目昆虫,双翅目昆虫,或鳞翅目害虫。
在另一优选例中,所述的黄烷酮3-羟化酶选自:
(a)如seqidno:2所示氨基酸序列的多肽;
(b)将seqidno:2所示氨基酸序列经过一个或多个(如1-30个,较佳地1-20个,更佳地1-10个,如1-5个,1-3个)氨基酸残基的取代、缺失或添加而形成的,且具有(a)多肽功能的由(a)衍生的蛋白;或
(c)与(a)限定的蛋白序列有80%以上(较佳地90%以上,更佳地95%以上,如98%或99%以上)同源性且具有(a)蛋白功能的由(a)衍生的蛋白。
在另一优选例中,所述的黄烷酮3-羟化酶编码基因是如下序列或其变体:seqidno:1。
在本发明的另一方面,提供一种黄烷酮3-羟化酶或其编码基因的用途,用于提高植物对抗昆虫的能力,或提高植物的花青素含量。
在本发明的另一方面,提供黄烷酮3-羟化酶或其编码基因的下调剂的用途,用于降低植物对抗昆虫的能力,或降低植物的花青素含量;较佳地,所述的黄烷酮3-羟化酶或其编码基因的下调剂是特异性干扰黄烷酮3-羟化酶基因表达的干扰分子。
在本发明的另一方面,提供一种黄烷酮3-羟化酶或其编码基因的用途,用于作为鉴定植物对抗昆虫能力高低的分子标记,或用于作为鉴定植物花青素含量高低的分子标记。
在一个优选例中,若经检测植物组织中黄烷酮3-羟化酶的表达高于一个特定值,则相对而言,所述植物的抗昆虫能力高;若经检测植物组织中黄烷酮3-羟化酶表达低于一个特定值,则相对而言,所述植物的抗昆虫能力低。其中,除非另外说明,所述的“特定值”是指特定植物中黄烷酮3-羟化酶的表达量的平均值。
本发明的其它方面由于本文的公开内容,对本领域的技术人员而言是显而易见的。
附图说明
图1、osf3h转基因植株根切片。
a:osf3h基因的t0代超表达转基因植株的real-time定量表达检测;
b:osf3h基因的t0代rnai转基因植株的real-time定量表达检测;
c:osf3h-oe转基因植株根的横切片;
d:osf3h-rnai转基因植株根的横切片;
e:野生型zh11植株根的横切片。
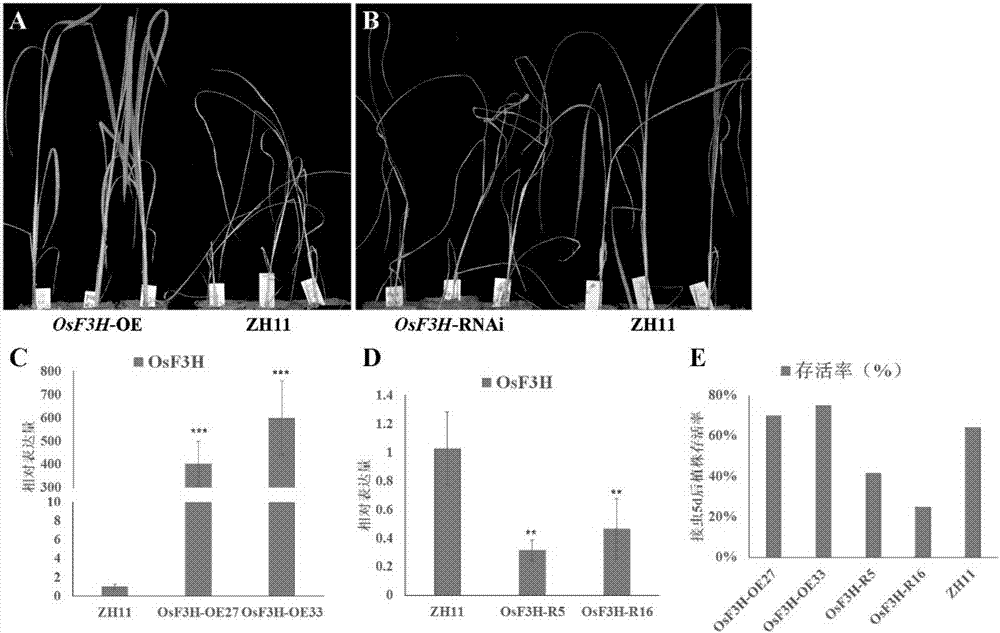
图2、osf3h转基因植株抗昆虫能力鉴定
a:t2代超表达植株抗昆虫能力鉴定;
b:t2代rnai植株抗昆虫能力鉴定;
c:t2代超表达植株osf3h基因的定量表达检测;
d:t2代rnai植株中osf3h基因的定量表达检测;
e:osf3h转基因植株t2代抗昆虫能力鉴定结果统计。
图3、拟南芥osf3h-oe转基因植株表型。
a:拟南芥野生型植株表型;
b:拟南芥osf3h-oe转基因植株t1代表型;
c:提取的花青素溶液的外观颜色;
d:拟南芥osf3h-oe转基因植株t2代花青素含量。
图4、osf3h基因编码蛋白的亚细胞定位。
图5、超表达载体p130135s-nos的图谱。
图6、p1301-rnai载体的图谱。
具体实施方式
本发明人在筛选植物抗虫基因的过程中,发现黄烷酮3-羟化酶(f3h)与植物对昆虫的抗性密切相关,促进黄烷酮3-羟化酶的表达可赋予植物对于昆虫的抗性,而下调其表达使得植物对昆虫更为敏感,更易于受到侵害。并且,促进黄烷酮3-羟化酶的表达还可提高植物的花青素含量。因此,黄烷酮3-羟化酶(或其编码基因)或调节黄烷酮3-羟化酶(或其编码基因)的物质和方法可应用于植物育种中,选育出抗虫性能发生变化的植物品种,或花青素含量发生变化的植物品种。
如本文所用,所述的“植物(作物)”包括但不限于:十字花科植物,禾本科植物、茄科植物、大戟科植物等。所述的植物可以是:双子叶植物、单子叶植物、或裸子植物。比如,所述的“植物”包括但不限于:水稻、小麦、玉米、马铃薯、木薯等。较佳的,所述的植物是禾本科植物或十字花科植物。
本发明中,所述的有害昆虫包括:飞虱科昆虫,同翅目昆虫,双翅目昆虫鳞翅目昆虫(如二化螟)等。
如本文所用,所述的“飞虱科昆虫”包括但不限于:褐飞虱(brownplanthopper,bph),白背飞虱(whitebackplanthopper,wbph)及灰飞虱(smallbrownplanthopper,sbph)。“飞虱科”是属于同翅目的1个科,“飞虱科昆虫”具有较多共性如下:通称飞虱,全部植食性,很多种生活于禾本科植物,为刺吸式农业害虫。有迁飞习性,是我国和许多亚洲国家当前水稻上的首要害虫。其中褐飞虱为单食性害虫,在水稻和普通野生稻上取食和繁殖后代。白背飞虱和灰飞虱食性广,可危害水稻、小麦、玉米等禾本科植物。
本发明还包括各黄烷酮3-羟化酶的片段、衍生物和类似物。如本文所用,术语“片段”、“衍生物”和“类似物”是指基本上保持本发明的黄烷酮3-羟化酶相同的生物学功能或活性的多肽。本发明的多肽片段、衍生物或类似物可以是(i)有一个或多个(如1-30个;较佳地1-20个;更佳地1-10个;如5个,3个)保守或非保守性氨基酸残基(优选保守性氨基酸残基)被取代的蛋白,而这样的取代的氨基酸残基可以是也可以不是由遗传密码编码的,或(ii)在一个或多个(如1-30个;较佳地1-20个;更佳地1-10个;如5个,3个)氨基酸残基中具有取代基团的蛋白,或(iii)附加的氨基酸序列融合到此蛋白序列而形成的蛋白等。根据本文的定义这些片段、衍生物和类似物属于本领域熟练技术人员公知的范围。例如,在本领域中,用性能相近或相似的氨基酸进行取代时,通常不会改变蛋白质的功能。又比如,在c末端和/或n末端添加或缺失一个或数个氨基酸通常也不会改变蛋白质的功能。该术语还包括黄烷酮3-羟化酶的活性片段和活性衍生物。
任何一种黄烷酮3-羟化酶的生物活性片段都可以应用到本发明中。在这里,黄烷酮3-羟化酶的生物活性片段的含义是指作为一种蛋白,其仍然能保持全长的黄烷酮3-羟化酶的全部或部分功能。通常情况下,所述的生物活性片段至少保持50%的全长黄烷酮3-羟化酶的活性。在更优选的条件下,所述活性片段能够保持全长黄烷酮3-羟化酶的60%、70%、80%、90%、95%、99%、或100%的活性。
编码黄烷酮3-羟化酶或其保守性变异蛋白的多核苷酸序列(编码序列)也可以应用到本发明中。编码成熟黄烷酮3-羟化酶的编码区序列可以与seqidno:1所示的序列基本上相同或者是简并的变异体。如本文所用,“简并的变异体”在本发明中是指编码具有seqidno:2所示的多肽,但与seqidno:1所示的序列有差别的核酸序列。
术语“编码基因”可以是包括编码所述蛋白的多核苷酸,也可以是还包括附加编码和/或非编码序列的多核苷酸。
上述多核苷酸的变异体也是可用的,其编码与本发明有相同的氨基酸序列的蛋白或蛋白的片段、类似物和衍生物。此多核苷酸的变异体可以是天然发生的等位变异体或非天然发生的变异体。这些核苷酸变异体包括取代变异体、缺失变异体和插入变异体。如本领域所知的,等位变异体是一个多核苷酸的替换形式,它可能是一个或多个核苷酸的取代、缺失或插入,但不会从实质上改变其编码的蛋白的功能。
应理解,虽然本发明的黄烷酮3-羟化酶编码基因(f3h)优选获自禾本科植物,但是获自其它植物的与该f3h基因高度同源(如具有80%以上,如85%、90%、95%、甚至98%序列相同性)的其它基因也在本发明考虑的范围之内。比对序列相同性的方法和工具也是本领域周知的,例如blast。本领域已知,f3h基因的cdna最初从金鱼草中克隆得到,已经在多种植物中克隆得到,如苹果、紫花苜蓿、玉米和银杏等,其作用底物是柚皮素,说明这种基因保守存在于很多物种中。
本发明的f3h通常可以用pcr扩增法、重组法或人工合成的方法获得。对于pcr扩增法,可根据本发明所公开的有关核苷酸序列,尤其是开放阅读框序列来设计引物,并用市售的cdna库或按本领域技术人员已知的常规方法所制备的cdna库作为模板,扩增而得有关序列。此外,还可用人工合成的方法来合成有关序列。
包含所述编码序列的载体,以及用所述的载体或黄烷酮3-羟化酶编码序列经基因工程产生的宿主细胞也包括在本发明中。本领域的技术人员熟知的方法能用于构建含黄烷酮3-羟化酶编码序列和合适的转录/翻译控制信号的表达载体。这些方法包括体外重组dna技术、dna合成技术、体内重组技术等。所述的序列可有效连接到表达载体中的适当启动子上,以指导mrna合成。包含上述的适当编码序列以及适当启动子或者控制序列的载体,可以用于转化适当的宿主细胞,以使其能够表达蛋白质。
宿主细胞通常是植物细胞。转化植物一般可使用农杆菌转化或基因枪转化等方法,例如叶盘法、幼胚转化法等;优选的是农杆菌法。对于转化的植物细胞、组织或器官可以用常规方法再生成植株,从而获得相对于野生型而言性状发生改变的植物。
基于本发明人的新发现,本发明提供了所述的黄烷酮3-羟化酶或其编码基因的用途,用于调节(包括上调或下调)植物对抗昆虫的能力或调节植物花青素含量。在一种方式下,过表达正义黄烷酮3-羟化酶可以提高植物对抗昆虫的能力,或提高植物的花青素含量。在另一种方式下,下调黄烷酮3-羟化酶表达或活性后,可降低植物对抗昆虫的能力,或降低植物的花青素含量。本发明人发现,下调植株的感虫性状非常明显,在水稻基础研究中具有实际意义。因此,可基于黄烷酮3-羟化酶对于植物的影响作用来改变植物,从而达到根据实际生产需要改良植物品质的目的,例如制造具有强的抗虫能力的植物或花青素含量高的植物,或制造对于昆虫敏感的植物或花青素含量低的植物。
花青素是一类重要的水溶性天然植物色素,属于类黄酮物质,在植物体中广泛分布。苯丙氨酸代谢途径是花青素的生物合成途径。前体物质丙二酰辅酶a和香豆酰辅酶a在查尔酮合成酶(chs)和查尔酮异构酶(chi)的催化下形成无色的柚皮素,柚皮素由黄烷酮3-羟化酶(f3h)催化成二氢山奈素(dhk),dhk可以在f3’h和f3’5’h的催化下形成二氢栎皮黄酮(dhq)和二氢杨梅黄酮(dhm),f3’5’h也能将dhq催化成dhm。无色的dhk、dhq和dhm在二羟基黄酮醇还原酶(dfr)的催化下会进一步还原成不稳定的无色花色素,然后无色花色素在花色素苷合成酶(ans)和类黄酮3-o-糖基转移酶(ufgt)的作用下合成各种花色素苷进而合成各种花青素。
本发明还涉及黄烷酮3-羟化酶或其编码基因的上调剂或下调剂(如反义的f3h基因或如mirna)及其用途。由于上调剂或下调剂可调节f3h的表达和/或调节f3h的活性等,因此,所述的上调剂或下调剂可通过对f3h的影响来调节植物性状,从而达到改良植物的目的。
任何可提高黄烷酮3-羟化酶的活性、提高黄烷酮3-羟化酶的稳定性、促进黄烷酮3-羟化酶的表达、延长黄烷酮3-羟化酶有效作用时间、或促进f3h基因的转录和翻译的物质均可用于本发明,作为可用于提高植物对抗昆虫的能力,或提高植物的花青素含量的有效物质。
任何可降低黄烷酮3-羟化酶的活性、降低黄烷酮3-羟化酶的稳定性、抑制黄烷酮3-羟化酶的表达、减少黄烷酮3-羟化酶有效作用时间、或降低f3h基因的转录和翻译的物质均可用于本发明,作为可用于降低植物对抗昆虫的能力,或降低植物的花青素含量的有效物质。
本发明还涉及一种改良植物的方法,该方法包括调节所述植物中黄烷酮3-羟化酶的表达或活性。
在得知了所述的黄烷酮3-羟化酶的用途后,可以采用本领域人员熟知的多种方法来调节所述的黄烷酮3-羟化酶的表达。比如可通过一定的途径将携带f3h基因的表达单位(比如表达载体或病毒等)递送到靶点上,并使之表达活性的黄烷酮3-羟化酶。此外,也可以采用本领域人员熟知的多种方法来降低黄烷酮3-羟化酶的表达或使之缺失表达,比如将携带反义f3h基因的表达单位(比如表达载体或病毒等)递送到靶点上,使得细胞或植物组织不表达或降低表达黄烷酮3-羟化酶;或将f3h基因进行敲除。
作为一种可实施方式,将黄烷酮3-羟化酶的编码基因通过常规的方法克隆到适当的载体中,将所述的带有外源基因的重组载体导入到可表达所述黄烷酮3-羟化酶的植物细胞中,使所述的植物细胞表达黄烷酮3-羟化酶。可通过将所述植物细胞再生成植物,获得过量表达黄烷酮3-羟化酶的植物。优选的,利用农杆菌转化法将黄烷酮3-羟化酶的编码基因或反义基因转入植物中。
可采用任何适当的常规手段,包括试剂、温度、压力条件等来实施所述的方法。其它调节黄烷酮3-羟化酶表达的方法是本领域周知的。例如,可通过用强启动子驱动从而增强黄烷酮3-羟化酶的表达。或者通过增强子来增强该黄烷酮3-羟化酶基因的表达。适用于本发明方法的强启动子包括但不限于:35s启动子、水稻、玉米的ubi启动子等。
此外,本发明还涉及利用黄烷酮3-羟化酶或其编码基因作为一种基因转化植株后代的追踪标记。本发明还涉及利用黄烷酮3-羟化酶或其编码基因作为一种分子标记,通过检测植物中黄烷酮3-羟化酶的表达情况,鉴定植物对抗昆虫能力高低的分子标记,或用于鉴定植物花青素含量高低。
下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件如j.萨姆布鲁克等编著,分子克隆实验指南,第三版,科学出版社,2002中所述的条件,或按照制造厂商所建议的条件。
材料与方法:
1、材料
用于本发明的水稻材料为粳稻品种(oryzasatival.subsp.japonica)中花11,种植于中科院上海生命科学研究院植物生理生态研究所人工气候室,其生长环境为温度29℃±1℃,湿度50-70%±5%,光照时间7:00-19:00。
2方法
2.1osf3h超表达载体和表达下调载体的构建
为了从水稻中克隆osf3h基因,本发明人根据数据库(http://rice.plantbiology.msu.edu/)中atg到tga的cdna全序列,设计了如下引物:
正向引物:5’-cgggatccgccaagttgtgtgtgtgtacc-3’(seqidno:3);
反向引物:5’-ggggtacccgagaaactacgggggtgtt-3’(seqidno:4);
引物中,引入的bamhi和kpni位点用下划线表示。
用trizol(购自invitrogen)抽提野生型水稻zh11五叶期叶片的总rna,经dnasei消化去除可能的dna污染后,反转录得到cdna,用该cdna为模板,通过pcr扩增得到osf3h基因的全长cdna序列,通过上述双酶切克隆到超表达载体p130135s-nos(图5)中。
根据osf3h基因特异序列设计如下引物:
正向引物:5’-gggactagtggtaccgagcgatatccgagagcctg-3’(seqidno:5);
反向引物:5’-gggagctcggatccttggtcgaggtttctgctcc-3’(seqidno:6)。
下划线表示酶切位点,分别是正向引物中的spei和kpni位点,反向引物中的saci和bamhi位点。
构建了rnai载体:以上述获得的osf3hcdna为模板,通过pcr扩增到特异片段,通过kpni和bamhi双酶切和saci和spei双酶切分别将正向片段和反向片段克隆到p1301-rnai载体(图6)。
2.2转化水稻的鉴定及检测
构建好的超表达和rnai载体通过农杆菌介导的方法对水稻幼胚进行转化。trizol法抽提转化植株和野生型zh11相同生长时期相同部位的总rna,利用试剂盒(购自toyobo公司)消化提纯后取2μg进行反转录。
real-timepcr的条件是第1步95℃,3min;第2步95℃,15s;第3步60℃,30s;2-3步骤40个循环。
利用引物来检测是否转化成功,引物序列如下:
正向引物5’-aggtggatcgccgtgaatc-3’(seqidno:7);
反向引物5’-gctcttgtatcttccgttgcttag-3’(seqidno:8)。
2.3转基因水稻切片观察
取转基因植株和zh11相同时期相同部位的根部横切组织5mm左右,用50%的faa固定液固定,真空离心机抽气15min后换新鲜固定液,4℃保存过夜,送本单位仪器中心进行树脂切片的制作。切片观察时取叶片横切面上的相同部位进行比较,观察是否出现细胞数目的变化。
2.4转基因水稻抗昆虫能力鉴定
以野生型zh11为对照,取35-40天(d)左右的转基因植株进行褐飞虱抗性的鉴定,每株苗接2龄-3龄的虫10头。接虫后第2天重新确认每株苗存活的虫量,5-6天后观察植株的死亡率。
2.4花青素测定
取相同时期相同部位的组织,液氮将其磨成粉末,加入花青素提取液中,4℃过夜,第二天4℃离心(16000g,10min)后取上清液观察测定。花青素提取液中,甲醇:hcl:h2o比例分别为85%,5%和15%。
2.5亚细胞定位
扩增得到osf3h基因的全长序列片段,以蛋白融合的方式克隆到含gfp的蛋白表达载体pa7上,设计如下引物用于扩增:
正向引物:5’-ccgctcgagatggcggaccagctcatctc-3’(seqidno:9);
反向引物:5’-ggactagtggagatgtgtctgtaggtgttg-3’(seqidno:10);
上述引物中,引入的xhoi和spei位点用下划线表示。
制备水稻原生质体后,将融合的质粒dna和pa7空载体分别取5-10μg与100μl原生质体混合,加入110μl新配制的pegsolution混合液(40%peg4000,0.2m甘露醇,0.1mcacl2),在室温、暗下放置10-20min。轻轻缓慢加入440μlw5溶液,轻轻翻转管子使之混匀,离心1500rpm,3min。收集原生质体,轻轻用1mlwi溶液(0.5m甘露醇,20mmkcl,4mmmesph5.7)重悬原生质体。最后,将原生质体转移至六孔板,在暗下室温放置6-16h后用于confocol观察。
实施例1、osf3h基因的克隆
osf3h基因的公共序列号为loc_03g03034(http://rice.plantbiology.msu.edu/)。其cdna序列如下(seqidno:1):atggcggaccagctcatctccacggcagaccacgacacgctgccgggcaactacgtgcgccccgaggcgcagcgcccgcgcctcgccgacgtgctctccgacgcctccatccccgtcgtcgacctcgccaaccccgaccgcgccaagctcgtctcccaggtcggcgccgcctgccgctcccacggcttcttccaggtgctcaaccatggggtgccagtggagctgacactgtcggtgctggcggtggcgcacgacttcttccggctgccggcggaggagaaggccaagctctactccgacgacccggccaagaagatccgcctctccaccagcttcaacgtccgcaaggagaccgtgcacaactggcgcgactacctccgcctccactgctacccgcttcaccgctacctccctgattggccatccaaccccccttccttcagggagatcataagcacatactgcaaagaagttcgggagctcggattcagactgtacggagcgatatccgagagcctgggcttggaacaggactacatcaagaaggttcttggtgagcaggagcagcatatggcggtgaacttctaccccaagtgcccggagccagagctgacgttcggactgccggcgcacaccgacccgaacgccctcaccatcctcctcatggaccagcaggtggccggcctgcaagttctcaaggaaggcaggtggatcgccgtgaatccacagcccaacgcgctggtgatcaacattggtgatcagctacaggcgctaagcaacggaagatacaagagcgtgtggcaccgtgctgtcgtcaactctgacaaagcgaggatgtccgtcgcatcgttcctgtgcccctgcaacgacgtgctcatcggcccagctcagaagctcatcaccgatggctccccggccgtctaccggaactacacctacgacgagtactacaagaagttctggagcagaaacctcgaccaagaacactgcttggagctcttcagaacaacacctacagacacatcttga
其编码的蛋白的氨基酸序列如下(seqidno:2):
madqlistadhdtlpgnyvrpeaqrprladvlsdasipvvdlanpdraklvsqvgaacrshgffqvlnhgvpveltlsvlavahdffrlpaeekaklysddpakkirlstsfnvrketvhnwrdylrlhcyplhrylpdwpsnppsfreiistyckevrelgfrlygaiseslgleqdyikkvlgeqeqhmavnfypkcpepeltfglpahtdpnaltillmdqqvaglqvlkegrwiavnpqpnalvinigdqlqalsngryksvwhravvnsdkarmsvasflcpcndvligpaqklitdgspavyrnytydeyykkfwsrnldqehclelfrttptdts
本发明中,本发明人克隆了osf3h基因的全长并构建到质粒p1301-35snos中获得超表达植株。
本发明人还选取osf3h基因的一个特异片段(即seqidno:1中第500-981位),构建了osf3h基因的rnai载体,并通过遗传转化获得转基因植株。
同时,本发明人还用基因的全长序列构建了pa7-osf3h-gfp质粒。
实施例2、osf3h基因超表达和rnai的表型
本发明人克隆了水稻osf3h基因,构建osf3h-oe和osf3h-rnai质粒,并将质粒遗传转化到野生型水稻zh11中。经real-time检测获得超表达的转基因植株和osf3h基因下调的转基因植株,如图1a和1b所示。
osf3h-rnai的转基因植株的根明显变粗,切片显示根变粗是由于薄壁细胞增多的缘故,如图1d与e对照。除此之外,osf3h-oe和osf3h-rnai转基因植株没有发现明显的肉眼可见的发育相关的表型变化。
实施例3、osf3h-oe和osf3h-rnai转基因植株的抗昆虫能力检测
以野生型zh11为对照,取35-40d左右的t2代阳性转基因植株进行褐飞虱抗性的鉴定,发现osf3h-oe转基因植株对褐飞虱呈现出明显的抗性,如图2a;而osf3h-rnai转基因植株对褐飞虱呈现出明显的感虫表型,如图2b所示。
根据real-time的结果,显示转基因植株抗性的强弱同osf3h基因的表达量高低呈明显的相关性,如图2c-e所示。
实施例4、osf3h-oe质粒转化拟南芥
osf3h基因编码黄烷酮3-羟化酶,是已知的花青素合成途径(苯丙氨酸代谢)中的关键酶,它调控黄酮与花青素苷产物的合成,是整个黄酮类化合物代谢途径的中枢位点。由于水稻中花青素含量变化导致的表型变化不易观察,且植物的f3h基因具有较高的同源性,所以本发明人将osf3h基因的超表达质粒转化拟南芥后进行表型观察。
结果发现,osf3h-oe转基因拟南芥茎秆和叶片上的颜色明显比野生型深,如图3b所示。
花青素含量测定也揭示出超表达转基因植株中的花青素含量显著提高,如图3c和3d所示。
实施例5、osf3h基因的亚细胞定位
据报道,植物中的f3h主要定位于细胞质中。本发明人通过pcr方法扩增得到osf3h基因的全长序列片段,以蛋白融合的方式克隆到含gfp的蛋白表达载体pa7上,并转化到水稻原生质体中,通过confocol检测融合蛋白的亚细胞定位情况。
结果显示,f3h-gfp融合蛋白的确定位在细胞质中(图4)。
在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本申请所附权利要求书所限定的范围。
- 还没有人留言评论。精彩留言会获得点赞!