一种饱和脂肪酸特异性脂肪酶突变体及其应用的制作方法
本发明涉及一种饱和脂肪酸特异性脂肪酶突变体及其应用,属于基因工程技术领域。
背景技术:
脂肪酸是油脂化工中最基础,使用最为广泛的原料之一。天然油脂水解是获得游离脂肪酸的重要途径之一。脂肪酸的生产技术主要有皂化法、水解法、蒸汽裂解法以及酶水解法,其中酶水解法由于具有反应条件温和、不饱和脂肪酸不易被氧化、副产物少的优势,成为脂肪酸制取方法的研究热点。不同类型、不同碳链长度及不同比例的脂肪酸具有不同的功能。短链脂肪酸可作为饲料添加剂,促进动物生长。中链脂肪酸广泛应用于医药、保健食用油及养殖业。长链不饱和脂肪酸在代谢中具有广泛的生理活性,用于健康保健。植物油脂中不饱和脂肪酸含量较高,如大豆油、菜籽油和油茶油中不饱和脂肪酸含量分别为84.34%、77.70%和81.43%。长链饱和脂肪酸如棕榈酸和硬脂酸,广泛存在于猪、牛和羊等动物油中,主要用于生产作为乳化剂的脂肪酸盐,并作为常用的塑料制品的稳定剂。动物油中各类饱和脂肪酸质量分数之和通常在60%以上,而且脂肪酸链长多为十六碳、十八碳,合计可达90%以上,含有十六个及十八个碳原子的脂肪酸经甲酯化后得到的甲酯产品与矿石柴油具有相近的碳原子数,同时在很多理化性质上都有相似性,因此可以利用动物油水解制备脂肪酸作为生物柴油的合成原料。
脂肪酶(triacylglycerolhydrolase,lipase,ec3.1.1.3)是一类甘油三酯水解酶,在油水界面催化酯键的水解或者合成,是“绿色”油脂加工制造的重要工业酶。动物油是制备长链饱和脂肪酸的重要来源,但脂肪酶对其的水解率不高,仅为60%左右(饲料工业杂志2008第12期p8-11)。其可能的原因是在于脂肪酶对饱和脂肪酸的底物特异性无法满足油脂加工行业的需求,若能够进一步提高脂肪酶对饱和脂肪酸的特异性,可以预期提高对动物油脂的水解效率。
技术实现要素:
为解决上述问题,本发明通过理性设计华根霉(rhizopuschinensis)脂肪酶(rcl)突变位点,获得了饱和脂肪酸特异性提高的突变酶,并且提高了对动物油的水解率。
本发明的第一个目的是提供一种脂肪酶突变体,所述脂肪酶突变体相对于具有seqidno:1所示氨基酸序列的脂肪酶,包括脂肪酶底物结合口袋的一个、两个或三个氨基酸残基的变化;所述变化增加口袋空间位阻;所述变化与脂肪酶对饱和脂肪酸的特异性相关。
在本发明的一种实施方式中,所述脂肪酶突变体是在氨基酸序列如seqidno.1所示的序列的基础上,将第286位的苏氨酸突变为谷氨酰胺,或者将第281位的异亮氨酸突变为苯丙氨酸,或者将第116位的丙氨酸突变为色氨酸的同时将第281位的异亮氨酸突变为苯丙氨酸。
在本发明的一种实施方式中,所述脂肪酶突变体的氨基酸序列如seqidno.2、seqidno.3或seqidno.4所示。
在本发明的一种实施方式中,第286位的苏氨酸突变为谷氨酰胺得到突变体t286q;第281位的异亮氨酸突变为苯丙氨酸得到的突变体为i281f;将第116位的丙氨酸突变为色氨酸的同时将第281位的异亮氨酸突变为苯丙氨酸得到的突变体为a116w/i281f。
本发明的第二个目的是提供编码所述脂肪酶突变体的基因序列。
本发明的第三个目的是提供携带所述基因序列的质粒或细胞。
在本发明的一种实施方式中,所述细胞为细菌、真菌或古菌。
本发明的第四个目的是提供所述脂肪酶突变体在食品、保健品、医药领域的应用。
在本发明的一种实施方式中,所述应用是利用所述脂肪酶突变体水解动物油脂。
在本发明的一种实施方式中,所述应用包括水解猪油、牛油、羊油、鸡油或鸭油。
本发明的有益效果:
本发明根据华根霉脂肪酶(rcl)结构分析,对其进行定点突变,获得了脂肪酸特异性改变的突变酶。相比野生型rcl,突变酶t286q、i281f和a116w/i281f由于增强了对饱和脂肪酸的特异性,其对猪油水解率分别由55%提高到了85%、90%和86%
附图说明:
图1为脂肪酶的脂肪酸链长特异性(c2:pnpc2、c4:pnpc4、c5:pnpc5、c8:pnpc8、c12:pnpc12、c14:pnpc14、c16:pnpc16);
图2为脂肪酶对大豆油中饱和脂肪酸和不饱和脂肪酸的水解特性;
图3为温度对脂肪酶活力和稳定性的影响,其中a是温度对活性的影响,b是温度对稳定性的影响;
图4为ph对脂肪酶活力和稳定性的影响,其中a是ph对活性的影响,b是ph对稳定性的影响;
图5为脂肪酶催化大豆油水解率。
具体实施方式
原料大豆油皂化值的测定参照gb/t5534-2008。
水解率=(av0-av)/(sv-av)×100%
式中:av0为水解后样品的酸值,mgkoh/g;av、sv分别为原料大豆油酸值和皂化值,mgkoh/g。
脂肪酶水解大豆油试验方法:每份样品称取5g大豆油,将一定量的50mm磷酸钾缓冲液加入50ml三角瓶中,超声使底物充分乳化;大豆油水解的反应条件设定为反应时间24h,水油质量比1:1,加酶量500u/g(油重),ph8.0,温度40℃,反应后加入10ml95%乙醇终止反应,测定水解产物的酸值。酸值测定方法参照gb/t5009.37-2003。
脂肪酸组成成分分析:用正己烷萃取反应后的油脂混合物,通过薄层层析tlc分离得到游离脂肪酸置于10ml具塞比色管中,加入1ml2%h2so4-甲醇溶液,80℃水浴30min,取出比色管,冷却至室温,加入2ml正己烷,充分混匀,加入饱和nacl溶液至瓶口,剧烈振摇,静置离心分层,收集上层正己烷相,加入适量的无水硫酸钠,氮吹浓缩进样分析。
色谱柱:db-wax(30m×250μm×0.25μm);进样量:1μl;分流比:50:1;进样口温度:225℃;载气:氮气,30ml·min-1;氢气,45ml·min-1;空气,450ml·min-1;程序升温:180℃保持1.5min,以10℃·min-1升到210℃,保持2min,然后以5℃·min-1升到220℃保持5min;检测器:火焰离子检测器(fid),温度为250℃。每个样品检测13.5min。
在安捷伦gc6890n气相色谱仪工作站上采用面积归一化法就算大豆油脂肪酸组分的百分含量。
脂肪酶水解活力测定方法:脂肪酶水解对硝基苯酚酯产生对硝基苯酚和脂肪酸,对硝基苯酚在水溶液中显黄色,在410nm有最大的光吸收,通过测定对硝基苯酚在410nm处的光吸收可测得脂肪酶的活力。酶活的定义为:一定反应条件下每分钟产生1μmol对硝基苯酚的酶量为一个脂肪酶水解酶活国际单位。
实施例1:华根霉脂肪酶的定点突变
根据华根霉脂肪酶晶体结构分析和类似结构脂肪酶晶体结构比较,利用全质粒pcr技术进行定点突变和组合突变。通过对华根霉脂肪酶底物结合口袋定点进行定点突变。
表1突变位点所需的引物
酶底物结合口袋中氨基酸的亲疏水性以及侧链基团大小等因素可能会影响脂肪酶的底物特异性。因此,从增加口袋亲水性的角度设计三个突变体,包括l285q、t286q以及在序列h284和l285中间插入一个亲水氨基酸q,突变体命名为hql;从增加空间位阻的角度设计三个突变位点,包括a116w、i281f、a116w/i281f。
实施例2:突变酶的脂肪酸特异性研究
以不同烷基碳链长度的对硝基苯酚脂肪酸酯为底物(pnpc2、pnpc4、pnpc5、pnpc8、pnpc12、pnpc14、pnpc16),在ph8.0和温度为40℃下检测实施例1突变酶的脂肪酸链长特异性。
由图1可知,相较于野生型脂肪酶而言,突变酶hql对长链脂肪酸的特异性增强,其中对pnpc16的特异性最高,水解活力提高至野生型的2.72倍。突变酶a116w和l285q均对pnpc16的特异性最高,水解活性分别是野生型的1.23倍和1.50倍。突变酶i281f拓宽了水解脂肪酸链长范围,增加了对pnpc2的水解活性,对pnpc12的特异性最高,水解活性是野生型的2倍。另外,突变酶l285q、t286q、hql、a116w、a116w/i281f无法水解pnpc4,并且所有的突变酶对pnpc8的水解活性均降低了。通过对华根霉脂肪酶底物结合口袋定点突变我们获得了一系列脂肪酸特异性改变的突变酶,尤其是获得了对长链脂肪酸特异性增强的突变酶。
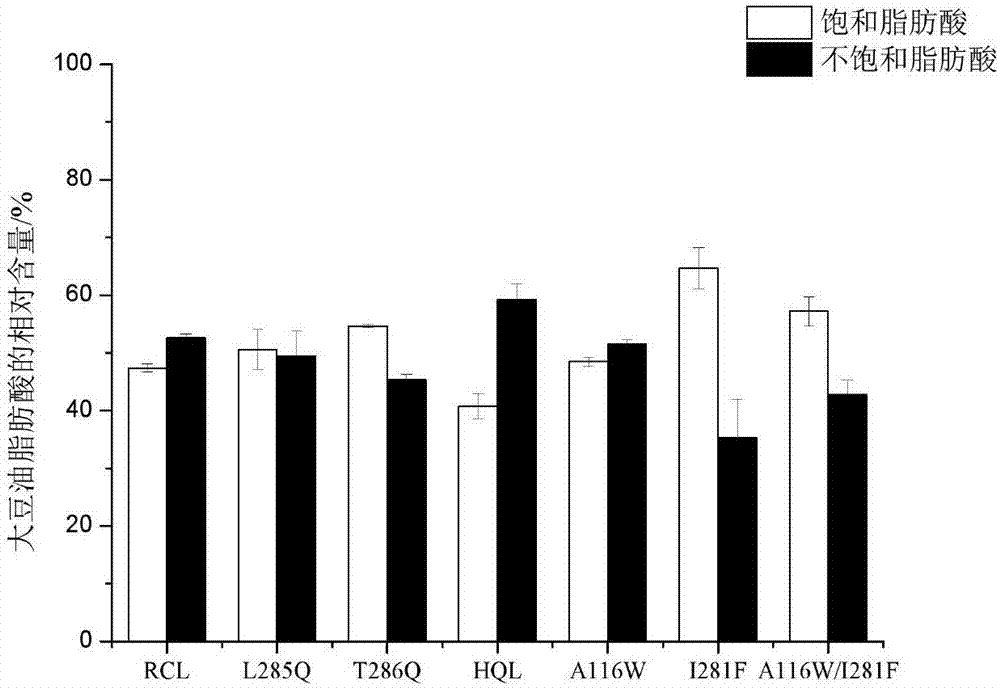
实施例3:突变酶对大豆油中脂肪酸的水解特性研究
为了考察脂肪酶对饱和及不饱和长链脂肪酸的底物特异性,以富含该类甘油三酯的大豆油作为底物进行测定。首先测定了大豆油的脂肪酸组成,含有13.77%饱和脂肪酸(棕榈酸11.19%,硬脂酸2.58%),以及86.23%不饱和脂肪酸(油酸25.21%,亚油酸54.67%,亚麻酸6.34%)。由表2和图2可知,突变酶hql增强了对不饱和脂肪酸的特异性,水解不饱和脂肪酸的活力是饱和脂肪酸的1.45倍,而野生型水解不饱和脂肪酸仅是饱和脂肪酸的1.10倍。突变酶hql对不饱和脂肪酸特异性更强,因此,可以预期hql对大豆油的水解率将会高于野生型酶。另外,突变酶t286q、i281f和a116w/i281f增强了对饱和脂肪酸的特异性,水解饱和脂肪酸的活力分别是不饱和脂肪酸的1.20、1.83和1.34倍,这几种脂肪酶突变体将更加适用于饱和脂肪酸含量高的动物油脂。
表2突变酶催化大豆油水解
实施例4:突变酶的热稳定性研究
以对硝基苯酚棕榈酸酯为底物(pnpc16)检测脂肪酶的活力。在不同的温度(20℃-60℃)下按标准方法测定酶液的脂肪酶活力,以最高酶活为相对酶活100%,研究温度对突变酶活力的影响。经过不同温度(20℃-60℃)下保温1h后测定酶液的脂肪酶活力,不保温的酶活力作为对照计算残余酶活力,考察温度对突变酶稳定性的影响。
由图3a可知所有突变酶催化反应的最适温度均为40℃,与野生型的最适温度一致。在20-40℃范围内,突变酶t286q的活力能够保持在84%以上,其他突变酶在20℃时均能保留65%以上的活力。但是当温度高于40℃后,酶蛋白逐渐变性,所有突变酶的活力均显著下降,当温度升至60℃时,突变酶hql的活力仍保留在20%左右,其他突变酶的活力均下降至10%以下。
图3b所示为脂肪酶的温度稳定性,在45℃时,突变酶hql和i281f的活力能保持在80%左右,只有突变酶a116w的活力下降至50%左右,而其他突变酶活力则保留在69%-75%之间。当温度超过55℃时,突变酶活力快速下降。
实施例5:突变酶的ph稳定性研究
ph影响酶的构象,也影响与催化有关基团的解离状况及底物分子的解离状态。酶的活力受环境ph的影响,在一定ph下,酶表现最大活力,高于或低于此ph,酶活力降低。分别配制0.05mol/l的磷酸盐缓冲液(ph6.5-ph8)、0.05mol/l的tris-hcl缓冲液(ph8-ph9)、0.05mol/l的碳酸盐缓冲(ph9-ph10),将酶液分别加在上述不同ph的缓冲液中,在标准条件下测定脂肪酶活力,以最高酶活为相对酶活100%,研究ph对突变酶活力的影响。酶液在上述不同ph的缓冲液中于25℃保温1h后测定酶液的脂肪酶活力,不保温的酶活力作为对照计算残余酶活力,考察ph对突变酶稳定性的影响。
由图4a可知所有突变酶催化反应的最适ph值均为8.0,与野生型的最适ph一致。ph范围在ph7.5-9.0之间,只有突变酶hqlt和i28f活力能保持在50%以上,其他突变酶在ph9.0时活力均低于50%。ph<7.5和ph>9的情况下,酶活力下降明显,说明反应ph显著影响突变酶的活力。
如图4b所示,所有突变酶均在最适ph8.0下稳定性最好。突变酶t286q相对于野生型稳定性较差,在ph7.5-8.5之间酶活力才能保持在60%以上,而其他突变酶在ph7.0和ph9.0时也能保留50%以上的活力。在ph<7和ph>9的情况下,稳定性极速减弱,只有突变酶i281f在ph10.0时还能保持22%的活力。
实施例6:不同突变酶催化大豆油水解能力比较
大豆油水解的反应条件设定为反应时间24h,水油质量比1:1,加酶量500u/g(油重),ph8.0,温度40℃,考察突变酶催化大豆油水解的能力。由图5可知,突变酶l285q与野生型催化大豆油的水解率接近,水解率为82%。突变酶a116w、i281f、a116w/i281f的催化大豆油水解的能力弱于野生型,水解率低至70%左右。突变酶l285q水解能力略高于野生型,水解率达到88%。突变酶hql催化大豆油水解的能力最高,达到98%左右。由于大豆油中不饱和脂肪酸含量高,占80%以上,而突变酶hql对长链不饱和脂肪酸特异性相比长链饱和脂肪酸特异性更强,因此催化水解大豆油的能力显著提高。
实施例7:不同突变酶催化猪油水解能力比较
猪油水解的反应条件设定为反应时间24h,水油质量比1:1,加酶量500u/g(油重),ph8.0,温度40℃,考察突变酶催化猪油水解的能力。结果表明,相比野生型rcl,突变酶t286q、i281f和a116w/i281f由于增强了对饱和脂肪酸的特异性,其对猪油水解率分别由55%提高到了85%、90%和86%。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动和修饰,因此本发明的保护范围应该以权力要求书所界定的为准。
序列表
<110>江南大学
<120>一种饱和脂肪酸特异性脂肪酶突变体及其应用
<160>14
<170>patentinversion3.3
<210>1
<211>296
<212>prt
<213>人工序列
<400>1
aspthrgluthrvalglyglymetthrleuaspleuprogluasnpro
151015
proproileproalathrserthralaproserseraspserglyglu
202530
valvalthralathralaalaglnilelysgluleuthrasntyrala
354045
glyvalalaalathralatyrcysargservalvalproglythrlys
505560
trpaspcyslysglncysleulystyrvalproaspglylysleuile
65707580
lysthrphethrserleuleuthraspthrasnglypheileleuarg
859095
seraspalaglnlysthriletyrvalthrpheargglythrasnser
100105110
pheargseralailethraspmetvalphethrphethrasptyrser
115120125
provallysglyalalysvalhisalaglypheleusersertyrasn
130135140
glnvalvallysasptyrpheprovalvalglnaspglnleuthrala
145150155160
tyrproasptyrlysvalilevalthrglyhisserleuglyglyala
165170175
glnalaleuleualaglymetaspleutyrglnargglulysargleu
180185190
serprolysasnleuseriletyrthrvalglycysproargvalgly
195200205
asnasnalaphealatyrtyrvalaspserthrglyileprophehis
210215220
argthrvalhislysargaspilevalprohisvalproproglnala
225230235240
pheglytyrleuhisproglyvalglusertrpilelysgluasppro
245250255
alaaspvalglnilecysthrserasnilegluthrlysglncysser
260265270
asnserilevalprophethrserilealaasphisleuthrtyrphe
275280285
glyileasngluglysercysleu
290295
<210>2
<211>296
<212>prt
<213>人工序列
<400>2
aspthrgluthrvalglyglymetthrleuaspleuprogluasnpro
151015
proproileproalathrserthralaproserseraspserglyglu
202530
valvalthralathralaalaglnilelysgluleuthrasntyrala
354045
glyvalalaalathralatyrcysargservalvalproglythrlys
505560
trpaspcyslysglncysleulystyrvalproaspglylysleuile
65707580
lysthrphethrserleuleuthraspthrasnglypheileleuarg
859095
seraspalaglnlysthriletyrvalthrpheargglythrasnser
100105110
pheargseralailethraspmetvalphethrphethrasptyrser
115120125
provallysglyalalysvalhisalaglypheleusersertyrasn
130135140
glnvalvallysasptyrpheprovalvalglnaspglnleuthrala
145150155160
tyrproasptyrlysvalilevalthrglyhisserleuglyglyala
165170175
glnalaleuleualaglymetaspleutyrglnargglulysargleu
180185190
serprolysasnleuseriletyrthrvalglycysproargvalgly
195200205
asnasnalaphealatyrtyrvalaspserthrglyileprophehis
210215220
argthrvalhislysargaspilevalprohisvalproproglnala
225230235240
pheglytyrleuhisproglyvalglusertrpilelysgluasppro
245250255
alaaspvalglnilecysthrserasnilegluthrlysglncysser
260265270
asnserilevalprophethrserilealaasphisleuglntyrphe
275280285
glyileasngluglysercysleu
290295
<210>3
<211>296
<212>prt
<213>人工序列
<400>3
aspthrgluthrvalglyglymetthrleuaspleuprogluasnpro
151015
proproileproalathrserthralaproserseraspserglyglu
202530
valvalthralathralaalaglnilelysgluleuthrasntyrala
354045
glyvalalaalathralatyrcysargservalvalproglythrlys
505560
trpaspcyslysglncysleulystyrvalproaspglylysleuile
65707580
lysthrphethrserleuleuthraspthrasnglypheileleuarg
859095
seraspalaglnlysthriletyrvalthrpheargglythrasnser
100105110
pheargseralailethraspmetvalphethrphethrasptyrser
115120125
provallysglyalalysvalhisalaglypheleusersertyrasn
130135140
glnvalvallysasptyrpheprovalvalglnaspglnleuthrala
145150155160
tyrproasptyrlysvalilevalthrglyhisserleuglyglyala
165170175
glnalaleuleualaglymetaspleutyrglnargglulysargleu
180185190
serprolysasnleuseriletyrthrvalglycysproargvalgly
195200205
asnasnalaphealatyrtyrvalaspserthrglyileprophehis
210215220
argthrvalhislysargaspilevalprohisvalproproglnala
225230235240
pheglytyrleuhisproglyvalglusertrpilelysgluasppro
245250255
alaaspvalglnilecysthrserasnilegluthrlysglncysser
260265270
asnserilevalprophethrserphealaasphisleuthrtyrphe
275280285
glyileasngluglysercysleu
290295
<210>4
<211>296
<212>prt
<213>人工序列
<400>4
aspthrgluthrvalglyglymetthrleuaspleuprogluasnpro
151015
proproileproalathrserthralaproserseraspserglyglu
202530
valvalthralathralaalaglnilelysgluleuthrasntyrala
354045
glyvalalaalathralatyrcysargservalvalproglythrlys
505560
trpaspcyslysglncysleulystyrvalproaspglylysleuile
65707580
lysthrphethrserleuleuthraspthrasnglypheileleuarg
859095
seraspalaglnlysthriletyrvalthrpheargglythrasnser
100105110
pheargsertrpilethraspmetvalphethrphethrasptyrser
115120125
provallysglyalalysvalhisalaglypheleusersertyrasn
130135140
glnvalvallysasptyrpheprovalvalglnaspglnleuthrala
145150155160
tyrproasptyrlysvalilevalthrglyhisserleuglyglyala
165170175
glnalaleuleualaglymetaspleutyrglnargglulysargleu
180185190
serprolysasnleuseriletyrthrvalglycysproargvalgly
195200205
asnasnalaphealatyrtyrvalaspserthrglyileprophehis
210215220
argthrvalhislysargaspilevalprohisvalproproglnala
225230235240
pheglytyrleuhisproglyvalglusertrpilelysgluasppro
245250255
alaaspvalglnilecysthrserasnilegluthrlysglncysser
260265270
asnserilevalprophethrserphealaasphisleuthrtyrphe
275280285
glyileasngluglysercysleu
290295
<210>5
<211>44
<212>dna
<213>人工序列
<400>5
tttcacctctatcgctgatcaccaaacctactttggtattaacg44
<210>6
<211>44
<212>dna
<213>人工序列
<400>6
cgttaataccaaagtaggtttggtgatcagcgatagaggtgaaa44
<210>7
<211>49
<212>dna
<213>人工序列
<400>7
ctttcacctctatcgctgatcacttacaatactttggtattaacgaagg49
<210>8
<211>49
<212>dna
<213>人工序列
<400>8
ccttcgttaataccaaagtattgtaagtgatcagcgatagaggtgaaag49
<210>9
<211>45
<212>dna
<213>人工序列
<400>9
ttcacctctatcgctgatcaccaattaacctactttggtattaac45
<210>10
<211>45
<212>dna
<213>人工序列
<400>10
gttaataccaaagtaggttaattggtgatcagcgatagaggtgaa45
<210>11
<211>46
<212>dna
<213>人工序列
<400>11
gtggtactaattccttcagaagctggattactgacatggtcttcac46
<210>12
<211>46
<212>dna
<213>人工序列
<400>12
gtgaagaccatgtcagtaatccagcttctgaaggaattagtaccac46
<210>13
<211>44
<212>dna
<213>人工序列
<400>13
actctatcgttcctttcacctcttttgctgatcacttaacctac44
<210>14
<211>44
<212>dna
<213>人工序列
<400>14
gtaggttaagtgatcagcaaaagaggtgaaaggaacgatagagt44
- 还没有人留言评论。精彩留言会获得点赞!