抗流感病毒和金黄色葡萄球菌合并感染的新型抗感染策略的制作方法
本发明涉及用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染或单独细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂。本发明还提供包含用在预防和/或治疗包含细菌感染和流感病毒感染的合并感染或单独细菌感染中的这类抑制剂的组合物。此外,还提供一种体外测试系统,其中所述测试系统包含感染流感病毒和细菌或单独感染细菌的培养细胞。
背景技术:
流感a病毒是导致显著发病率和死亡率的严重呼吸道疾病的病原体。在感染流感病毒(iv)期间,大部分死亡案例实际上是由诸如金黄色葡萄球菌(s.aureus)、肺炎双球菌和流感嗜血杆菌的不同细菌引起的继发性肺炎所造成的(morens等人,2008,chertow等人,2013)。细菌合并感染最突出的问题是突然增强的致病性(lwao等人,2012,paddock等人,2012,parker等人,2012)和对抗不同病原体的有效抗感染药的有限的库存(arsenal)。流感病毒的高变异性和新菌株的不断出现(neumann等人,2009,taubenberger等人,2010,parry,2013)、菌株的具体特征(grundmann等人,2006,moran等人,2006,gillet等人,2007,shilo等人,2011)以及流感病毒(hayden等人,1992,bright等人,2006,pinto等人,2006,declercq等人,2007,pinto等人,2007)和细菌(grundmann等人,2006,moran等人,2006,shilo等人,2011)对可用的药物/抗体迅速产生的耐受性是造成治疗选择贫瘠的主要原因。此外,偶发的情况是,迄今为止治疗流感病毒和细菌合并感染不可能使用单一化合物。本发明解决这一问题在于其提出了通过使用单一药物对抗iv和金黄色葡萄球菌合并感染的新的抗感染策略。而且,本发明通过提供以细胞因子而非细菌本身为靶向的药物而解决了细菌迅速产生耐受性的问题。
发明概述
本发明的技术问题通过在权利要求中所反映的、在说明书中所描述的以及在实施例和附图中所列举的实施方案来解决。
如上所述,本发明涉及用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂。
此外,本发明涉及用在用于预防和/或治疗细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂。
尽管在上一世纪进行了密集的研究,iv仍然表现出对人类的严重威胁。对老年人和免疫功能低下的个体尤为危险的季节性暴发是由于感染流感a或b病毒。
在过去几十年内,耐甲氧西林金黄色葡萄球菌发病率提高,造成婴幼儿和儿童同时感染iv的问题(iverson等人,2011,thorburn等人,2012)。在细菌合并感染中出现的一个主要问题是可能由加速的细胞因子表达所引起的突然且高度增强的致病性,这导致组织损伤。特别地,在具有表达杀白细胞毒素(pvl)的金黄色葡萄球菌的合并感染中观察到严重的肺上皮细胞破坏,这是由于pvl-介导的中性粒细胞杀死后蛋白酶的失控性释放所致(gillet等人,2007,niemann等人,2012)。细菌合并感染通常发生在感染iv的前六天之内,这甚至会引发更多的爆发性疾病、肺炎以及更高的死亡率(iverson等人,2011,chertow等人,2013)。然而,在某些情况下,当病毒感染似乎已被清除时,细菌合并感染才发生。对于病毒/细菌合并感染的治疗,仅存在有限的可能性。
对抗流感的一种有前景的抗病毒策略基于以下事实:作为细胞内病原体的iv高度依赖于细胞信号传导机构(gong等人,2009,ludwig,2009)。iv获得为其自身目的抢夺细胞因子的能力(ludwig等人,2003)。而且,iv能够抑制其宿主的先天性免疫应答。考虑到这些依赖性,细胞病毒支持性功能是新型抗病毒干扰最具前景的候选者(ludwig等人,2003,ludwig,2011,planz,2013)。在过去的几年中,我们和其他人确定了raf/mek/erk促有丝分裂激酶级联(pleschka等人,2001,ludwig等人,2004,olschlager等人,2004,marjuki等人,2006,ludwig,2009,droebner等人,2011)、ikk/nfκb模块(pleschka等人,2001,wurzer等人,2004,marjuki等人,2006,mazur等人,2007,ludwig等人,2008,dudek等人,2010,droebner等人,2011,ehrhardt等人,2013,haasbach等人,2013)、p38-(borgeling等人,2014)以及pi3k-信号(ehrhardt等人,2006,ehrhardt等人,2007a,ehrhardt等人,2007b,ehrhardt等人,2009,eierhoff等人,2010)通路作为抗病毒方法的合适靶点。
以细胞而非病毒因子为靶向防止了耐受性问题,这是因为病原体并不能够替代丢失的细胞功能。对于一些细胞因子,化学化合物是有效的,尽管处于早期阶段,这些化学化合物中的一些已经进入临床试验甚至已经获得许可。
与iv复制相反,金黄色葡萄球菌门是与宿主细胞相独立的。新型抗菌替代品不以所述病原体产生的必需基因产品为靶点,而是在金黄色葡萄球菌感染期间不通过杀害细菌或增强宿主免疫力而抑制致病因子(park等人,2012)。其他策略防止金黄色葡萄球菌在人类宿主中的移植(park等人,2012)。这些化合物还显示出较低的引发耐受性的可能性。近期,越来越多的证据证明金黄色葡萄球菌在感染期间也会为其自身利益使用细胞信号(oviedo-boyso等人,2011),但这样的细菌-支持性细胞因子尚未被详细表征为抗菌疗法的靶点。
本发明惊奇地观察到,以前示出的具有抗流感活性的对抗细胞内信号因子的药物,诸如nfкb,mek或p38map激酶也表现出抗金黄色葡萄球菌活性,并且在合并感染情况下同时降低病毒和细菌的效价。因此,这些信号抑制剂是用于治疗单独的iv感染或金黄色葡萄球菌感染的最具前景的候选者,然而,最重要的是,其还对抗伴有细菌合并感染的严重流感。
在一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其中,所述细菌感染由以下细菌调解:葡萄球菌科、链球菌科、人菌科、假单胞菌科、衣原体科、支原体科、肠杆菌科、假单胞菌目和/或巴斯德氏菌科。
在另一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染的方法中,其中,所述流感病毒感染由流感a病毒或流感b病毒介导,优选地,所述流感a病毒为h1n1、h2n2、h3n2、h6n1、h7n7、h7n9、h9n2h10n7、h10n8或h5n1。在一个实施方案中,所述流感a病毒是h1n1。在其他实施方案中,所述流感a病毒为h3n2、h5n1和h7n9。在另外的实施方案中,所述流感a病毒为h3n2、h5n1、h1n1和h7n9。
在进一步的实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其中,所述mek抑制剂选自u0126、plx-4032、azd6244、azd8330、as-703026、gsk-1120212、rdea-119、ro-5126766、ro-4987655、ci-1040、pd-0325901、gdc-0973、tak-733、pd98059、arry-438162、pf-3644022和pd184352,优选为azd8330、gsk-1120212、u0126、gdc-0973、ci-1040、pd0325901、arry-438162、pf-3644022和azd6244,最优选为u0126、gdc-0973、ci-1040、azd8330和gsk-1120212。
在另一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其中,所述p38抑制剂选自sb202190、ly2228820、cay10571、sb203580、tie2激酶抑制剂、2-(4-氯苯基)-4-(氟苯基)-5-吡啶-4-基-1,2-二氢吡唑基-3-酮、cgh2466、sb220025、抗生素llz1640-2、tak715、sb202190盐酸盐、skf86002、amg548、cmpd-1、eo1428、jx401、ml3403、rwj67657、sb202190、sb203580、sb203580盐酸盐、sb239063、scio469、sx011、tak715、pamapimod、losmapimod(gw856553)、dilmapimod(sb681323)、vx702、vx745、doramapimod(birb796)、bms-582949、arry-797、ph797804,优选为vx-702、sb202190、pamapimod、losmapimod(gw856553)、dilmapimod(sb681323)、doramapimod(birb796)、bms-582949、arry-797、ph797804和scio-469。
在另一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其中,所述nfкb抑制剂选自lasag(也叫lg-asa)、sc75741、mg132、tpca-1、pctc、imd0354、木犀草素、咖啡酸苯乙酯、豆蔻明、pf184、ikk16、sc514、醉茄素a、牛蒡甙元、bay11-7085、psi、pr39、ro106-9920、bay11-7821、ml-130、南蛇藤醇、丹参酮iia、hu211、胶霉毒素、cid2858522、和厚朴酚、穿心莲内酯、10z-hymenialdisine、achp、扁塑藤素、柳氮磺胺吡啶、ml120b二盐酸盐、氨来占诺、9-甲基链霉戊二酰亚胺、n-硬脂酰基植物鞘氨醇、2-(1,8-二氮杂萘-2-基)-苯酚、5-氨基水杨酸、bay11-7085、3,4-二羟基肉桂酸乙酯、helanalin、nf-κb活化抑制剂ii、jsh-23、糖皮质激素受体调节器、cpda、ppm-18、阿司匹林(asa)、吡咯烷二硫代羧酸铵盐、(r)-mg132、sc75741楝酰胺、水杨酸钠、qnz、ps-1145、cay10512、硼替佐米、双水杨酯、白藜芦醇、脱氧精胍菌素、苏灵大、萨力多胺、agro-100、chs828和/或姜黄素,优选为硼替佐米、姜黄素、阿司匹林(asa)、双水杨酯、白藜芦醇、水杨酸钠、lasag(也叫lg-asa)、脱氧精胍菌素、苏灵大、萨力多胺、agro-100、chs828,更优选为sc75741、asa和lasag(也叫lg-asa),最优选为lasag(也叫lg-asa)。
在另外的实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其中,所述mek抑制剂与另外一种mek抑制剂、所述p38抑制剂和/或所述nfкb抑制剂联合;所述p38抑制剂与另外一种p38抑制剂、所述mek抑制剂和/或所述nfкb抑制剂联合;或者所述nfкb抑制剂与另外一种nfкb抑制剂、所述p38抑制剂和/或所述mek抑制剂联合。
在进一步的实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染的方法中,其中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂与针对所述流感病毒和/或所述细菌的一种或多种抑制剂联合。在一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂与针对所述流感病毒和/或所述细菌的一种或多种抑制剂同时施用、在针对所述流感病毒和/或所述细菌的一种或多种抑制剂之前或之后施用。这样,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂可以与1、2、3、4、5、6、7或8种针对流感病毒的抑制剂联合。类似地,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂可以与1、2、3、4、5、6、7或8种针对细菌的抑制剂联合。
在一个实施方案中,所述针对所述流感病毒的一种或多种抑制剂为神经氨酸苷酶抑制剂,优选为奥司他韦磷酸盐、扎那米韦、奥司他韦或帕拉米韦。
在另一个实施方案中,所述针对所述流感病毒的一种或多种抑制剂为针对离子通道蛋白(m2)的化合物,优选为金刚烷胺和/或金刚烷乙胺。
在进一步的实施方案中,所述针对所述流感病毒的一种或多种抑制剂为经由使用病毒聚合酶复合体成分、pb1、pb2、pa或np,优选为np阻断剂nucleozin或聚合酶抑制剂t-705干预聚合酶或核酸内切酶活性的化合物。
在进一步的实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的细菌感染的方法中,其中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂与针对所述细菌的一种或多种抑制剂联合。
在另一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其中,所述针对所述细菌的一种或多种抑制剂为抗生素,优选为庆大霉素、利福平、溶葡萄球菌酶(lysosthaphin)、红霉素、左氧氟沙星、替考拉宁、盘尼西林和苯甲异恶唑青霉素。
在另外的实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中,其用于患者,优选用于脊椎动物。
本发明还提供一种组合物,包含用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的方法中的mek抑制剂,p38抑制剂和/或nfкb抑制剂。优选地,所述组合物进一步包含载体。
本发明还涉及一种组合物,包含用在用于预防和/或治疗细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfкb抑制剂。优选地,所述组合物进一步包含载体。
本发明还提供一种组合物,包含用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的方法中的mek抑制剂,p38抑制剂和/或nfкb抑制剂以及针对所述流感病毒和/或所述细菌的一种或多种抑制剂。优选地,所述组合物进一步包含载体。
本发明还涉及一种组合物,包含用在用于预防和/或治疗细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfкb抑制剂以及针对所述细菌的一种或多种抑制剂的组合物。优选地,所述组合物进一步包含载体。
在进一步的实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的合并感染的方法中,其中,与接触前的体外测试系统相比,与体外测试系统相接触时,所述mek抑制剂,所述p38抑制剂和/或所述nfкb抑制剂同时降低了病毒和细菌感染,其中,所述测试系统包含感染
a)流感病毒,和
b)细菌的培养细胞。
在一个实施方案中,所述病毒感染的降低是指血小板形成单位(pfu)/ml的降低,所述细菌感染的降低是指菌落形成单位(cfu)/ml的降低。
在另一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfкb抑制剂用在用于预防和/或治疗本发明的细菌感染的方法中,其中,与接触前的体外测试系统相比,与体外测试系统相接触时,所述mek抑制剂,p38抑制剂和/或nfκb抑制剂会降低细菌感染,其中所述测试系统包括感染细菌的培养细胞。
本发明还涉及一种体外测试系统,其中所述测试系统包含感染
a)流感病毒,和
b)细菌的培养细胞。
本发明还提供本发明的所述体外测试系统用于确定抑制剂有效降低包含细菌感染和流感病毒感染的合并感染的用途。在一个实施方案中,所述病毒感染的降低是指血小板形成单位(pfu)/ml的降低,所述细菌感染的降低是指菌落形成单位(cfu)/ml的降低。
此外,本发明涉及一种检测有效预防和/或治疗包含细菌感染和流感病毒感染的合并感染的分子的方法,包括将感兴趣的化合物与本发明的体外测试系统接触,其中,与接触前的体外测试系统相比,所述感兴趣的化合物会同时降低病毒和细菌感染。
本发明还提供一种体外测试系统,其中所述体外测试系统包含感染细菌的培养细胞。
此外,本发明涉及本发明的体外测试系统用于确定有效降低细菌感染的抑制剂的用途。
此外,本发明涉及本发明的体外测试系统用于检查先天宿主细胞反应的用途,所述检查先天宿主细胞反应任选地包括检查信号传导水平、产生的细胞因子和趋化因子表达、细胞凋亡和坏死的诱导和/或调解健康和疾病的氧化还原止血作用。
本发明还提供一种检测有效预防和/或治疗细菌感染的分子的方法,包括将感兴趣的化合物与本发明的体外测试系统接触,其中,与接触前的体外测试系统相比,所述感兴趣的化合物会降低细菌感染。
此外,本发明涉及感染流感病毒和细菌的培养细胞。
本发明还提供感染细菌的培养细胞。
附图说明
附图显示:
图1:合并感染过程的时间标度。细胞感染iv30分钟。用金黄色葡萄球菌6850进行合并感染或细胞经空白处理(mock-treated)。细菌感染后3h细胞外细菌被溶解并通过抗生素处理移除。pbs清洗后,为细胞补充新鲜培养基(dmem/inv),病毒感染培养达18小时。
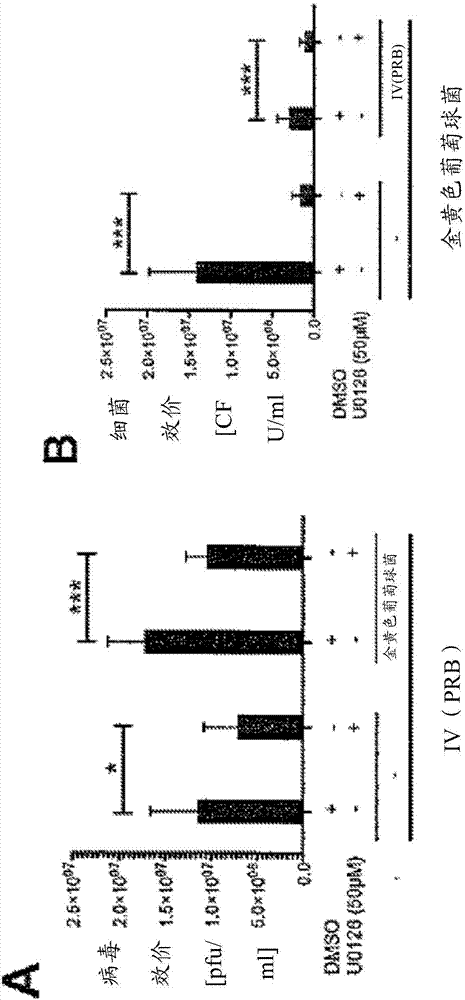
图2:mek抑制剂u0126降低iv效价(a/波多黎各/8/34)和金黄色葡萄球菌的负荷,即使在合并感染的情况下。在6孔培养板(8x105细胞/孔)上2mldmem[10%fcs]中接种人肺上皮细胞。接种16-20小时后,冲洗细胞并在37℃、感染复数(moi=0.1)下用pbs/ba[0.2%肽牛血清白蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素](每6孔500μl)或包含流感病毒a/波多黎各/8/34的pbs/ba进行培养。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞,并在50μmu0126或dmso存在下(溶剂对照)为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.5)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes](每6孔2ml)。细菌感染后3小时,用抗生素处理细胞以移除细胞外细菌。因此,用pbs清洗细胞并随后用mem/inv抗生素[2μg/ml溶葡球菌酶(sigma)](每6孔1ml)在37℃下培养20分钟。用pbs另外再清洗之后,为细胞补充包含50μmu0126或dmso以及0.333μg/ml胰蛋白酶(英杰公司)的dmem/inv。在37℃下进一步培养14小时后,如(hrincius等人,2010,tuchscherr等人,2011)中所述的确定iv效价和细胞内细菌。
iv效价描绘为血小板形成单位(pfu)/ml(a,c),金黄色葡萄球菌效价描绘为菌落形成单位(cfu)/ml(b,d)。数据代表两种生物样品三个独立实验的平均值±标准差(sd)。用双侧双样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。与用dmso处理的细菌相比,存在于50μmu0126中的金黄色葡萄球菌在培养18小时时导致降低的效价(e)。将限定数量的金黄色葡萄球菌6850悬浮培养液在补充有50μmu0126或dmso的dmem/inv中稀释,并在37℃下孵化18小时。稀释细菌,并通过在琼脂板上的连续稀释来确定细菌。数据代表两种生物样品三个独立实验的平均值±标准差。用双侧双样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。
图3:mek抑制剂u0126降低iv效价(a/fpv/布拉迪斯拉发/79)和金黄色葡萄球菌的负荷,即使在合并感染的情况下。在6孔培养板(8x105细胞/孔)上2mldmem[10%fcs]中接种人肺上皮细胞。接种16-20小时后,冲洗细胞并在37℃、感染复数(moi=0.001)下用pbs/ba[0.2%肽牛血清白蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素](每6孔500μl)或包含流感病毒a/fpv/布拉迪斯拉发/79的pbs/ba进行培养。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞,并在50μmu0126或dmso存在下(溶剂对照)为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.5)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes](每6孔2ml)。细菌感染后3小时,用抗生素处理细胞以移除细胞外细菌。因此,用pbs清洗细胞并随后用mem/inv抗生素[2μg/ml溶葡球菌酶(sigma)](每6孔1ml)在37℃下培养20分钟。用pbs另外再清洗之后,为细胞补充包含50μmu0126或dmso的dmem/inv。在37℃下进一步培养14小时后,如(hrincius等人,2010,tuchscherr等人,2011)中所述的确定iv效价和细胞内细菌。
iv效价描绘为血小板形成单位(pfu)/ml(a,c),金黄色葡萄球菌效价描绘为菌落形成单位(cfu)/ml(b,d)。数据代表两种生物样品三个独立实验的平均值±标准差(sd)。用双侧双样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。
图4:p38抑制剂sb202190降低iv的效价和金黄色葡萄球菌的负荷,即使在合并感染的情况下。在6孔培养板(8x105细胞/孔)上2mldmem[10%fcs]中接种人肺上皮细胞。接种16-20小时后,冲洗细胞并在37℃、感染复数(moi=0.1)下用pbs/ba[0.2%胎牛血清白蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素](每6孔500μl)或包含流感病毒a/波多黎各/8/34的pbs/ba进行培养。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞,并在10μmsb202190或dmso存在下(溶剂对照)为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.5)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes](每6孔2ml)。细菌感染后3小时,用抗生素处理细胞以移除细胞外细菌。因此,用pbs清洗细胞并随后用mem/inv抗生素[2μg/ml溶葡球菌酶(sigma)](每6孔1ml)在37℃下培养20分钟。用pbs另外再清洗之后,为细胞补充包含10μmsb202190或dmso以及0.333μg/ml胰蛋白酶(英杰公司)的dmem/inv。在37℃下进一步培养14小时后,如(hrincius等人,2010,tuchscherr等人,2011)中所述的确定iv效价和细胞内细菌。流感病毒效价描绘为血小板形成单位(pfu)/ml(a,c),金黄色葡萄球菌效价描绘为菌落形成单位(cfu)/ml(b,d)。数据代表两种生物样品三个独立实验的平均值±标准差(sd)。用双侧双样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。
图5:nf-kappab(nfκb)抑制剂lg-asa降低iv的效价和金黄色葡萄球菌的负荷,即使在合并感染的情况下。在6孔培养板(8x105细胞/孔)上2mldmem[10%fcs]中接种人肺上皮细胞。接种后16-20小时后,冲洗细胞并在37℃、感染复数(moi=0.1)下用pbs/ba[0.2%胎牛血清白蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素](每6孔500μl)或包含流感病毒a/波多黎各/8/34的pbs/ba进行培养。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞,并在5mmlg-asa存在下(溶剂对照)为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.5)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes](每6孔2ml)。细菌感染后3小时,用抗生素处理细胞以移除细胞外细菌。因此,用pbs清洗细胞并随后用dmem/inv抗生素[10%fbs,2μg/ml溶葡球菌酶(sigma)](每6孔1ml)在37℃下培养20分钟。用pbs另外再清洗之后,为细胞补充包含5mmlg-asa或水以及0.333μg/ml胰蛋白酶(英杰公司)的dmem/inv。在37℃下进一步培养14小时后,如(hrincius等人,2010,tuchscherr等人,2011)中所述的确定iv效价和细胞内细菌。流感病毒效价描绘为血小板形成单位(pfu)/ml(a,c),金黄色葡萄球菌效价描绘为菌落形成单位(cfu)/ml(b,d)。数据代表两种生物样品三个独立实验的平均值±标准差(sd)。用双侧双样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。
图6:病毒性神经氨酸苷酶抑制剂达菲降低iv的复制但加强金黄色葡萄球菌的负荷。在6孔培养板(8x105细胞/孔)上2mldmem[10%fcs]中接种人肺上皮细胞。接种后16-20小时,冲洗细胞并在37℃、感染复数(moi=0.001)下用pbs/ba[0.2%胎牛血清白蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素](每6孔500μl)或包含流感病毒a/fpv/布拉迪斯拉发/79的pbs/ba进行培养。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞,并在2μm达菲或羟乙基哌嗪乙磺酸(hepes)存在下(溶剂对照)为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.5)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes](每6孔2ml)。细菌感染后3小时,用抗生素处理细胞以移除细胞外细菌。因此,用pbs清洗细胞并随后用dmem/inv抗生素[2μg/ml溶葡球菌酶(sigma)](每6孔1ml)在37℃下培养20分钟。用pbs另外再清洗之后,为细胞补充包含2μm达菲或hepes的dmem/inv。在37℃下进一步培养14小时后,如(hrincius等人,2010,tuchscherr等人,2011)中所述的确定iv效价和细胞内细菌。
图7:经lg-asa处理会降低细胞内金黄色葡萄球菌6850的效价。在指定数量的lg-asa存在(a,c)和不存在(b,d)的情况下,用0.5moi金黄色葡萄球菌6850dmem/inv[1%人血清白蛋白,25nmol/ihepes]感染人肺上皮细胞(a549)3小时。感染后3小时,用dmem/inv抗生素[2μg/ml溶葡球菌酶(sigma)]进行抗生素清洗以移除非内化细菌,随后为细胞补充包含指定数量的lg-asa的dmem/inv。感染后18小时,通过光学显微镜观察细胞形态(a,b),通过在琼脂板上的连续稀释确定内化细菌的数量(c,d)。
图8:表2:p38抑制剂
图9:表3:nfκb抑制剂
图10:表4:nfκb抑制剂
图11:表5:抗生素
图12:体外合并感染过程的时间标度。在指定感染复数(moi)下用流感a病毒(iav)感染人肺上皮细胞(a549)30分钟,37℃下在pbs/ba[0.2%牛血清蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素]中溶解。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞。然后,用金黄色葡萄球菌6850进行细菌感染或将细胞进行空白处理。因此,除了指定数量的抑制剂(u0126或cl-1040)或溶剂对照以外,还为细胞补充含有或不含金黄色葡萄球菌的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes]。细菌感染后3小时,引入抗生素清洗[dmem,10%fbs,2μg/ml溶葡球菌酶或100μg/ml庆大霉素,20分钟]以移除非内化细菌。另外进行pbs清洗后,在存在或不存在所述抗体的情况下为细胞补充感染培养基dmem(0.2%ba,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素),37℃下病毒感染后培养达18小时。
图13:mek/erk信号传导的抑制会增强单一感染或合并感染后的细胞生存。
在37℃,感染复数(moi=0.01)(h7n7)或(moi=0.5)(h3n2)下,使人肺上皮细胞(a549)感染禽流感病毒株a/fpv/布拉迪斯拉发(bratislava)/79(h7n7)(fpv)或人流感病毒株a/威斯康星州(wisconsin)/67/2005(h3n2)。30分钟后,将病毒稀释物移除,用pbs清洗细胞,并在50μmu0126或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.1)的侵染培养基dmem/inv[包含1%人血清白蛋白,25nmhepes]。细菌感染后3小时,用包含10%fbs,2μg/ml溶葡球菌酶的dmem/fbs处理细胞20分钟以移除非内化细菌。用pbs另外再清洗后,为细胞补充包含50μmu0126或溶剂的感染培养基dmem/ba(0.2%ba,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素),37℃下培养18小时后,用光学显微镜观察细胞形态。
图14:mek/erk信号传导的抑制会引起单一感染或合并感染期间病毒效价的降低。
在37℃,感染复数(moi=0.01)(a,c),(moi=0.5)(b,d)下,使人肺上皮细胞(a549)感染禽流感病毒株a/fpv/布拉迪斯拉发/79(h7n7)(a,c)或人流感病毒株a/威斯康星州/67/2005(h3n2)(b,d)。30分钟后,将病毒稀释物移除,用pbs清洗细胞,并在50μmu0126或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.1)的侵染培养基dmem/inv[包含1%人血清白蛋白,25nmhepes]。细菌感染后3小时,用包含10%fbs,2μg/ml溶葡球菌酶的dmem/fbs处理细胞20分钟以移除细胞外细菌。另外进行pbs清洗后,为细胞补充包含50μmu0126或溶剂的感染培养基dmem/ba(0.2%ba,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素)。37℃下培养18小时后,通过标准空斑试验确定病毒效价。病毒效价以线性(a,b)或对数刻度(c,d)描绘为血小板形成单位/ml(pfu/ml)。数据代表两种生物样品四个独立实验的平均值±标准差(sd)。用双侧单样本t检验(*p<0.05;**p<0.01)来评估统计学显著性。
图15:通过施用u0126的mek抑制引起细菌生长的降低
在37℃,感染复数(moi=0.01)(h7n7)或(moi=0.5)(/h3n2)下,使人肺上皮细胞(a549)感染禽流感病毒株a/fpv/布拉迪斯拉发/79(h7n7)或人流感病毒a/威斯康星州/67/2005(h3n2)(a,c)。30分钟后,将病毒稀释物移除,用pbs清洗细胞,并在50μmu0126或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.1)(a)的侵染培养基dmem/inv(包含1%人血清白蛋白,25nmhepes)。细菌感染后3小时,用包含10%fbs,2μg/ml溶葡球菌酶的dmem/fbs处理细胞20分钟(a,c)以移除非内化细菌。另外进行pbs清洗后,为细胞补充包含50μmu0126或溶剂的感染培养基dmem/ba(0.2%ba,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素),感染18小时后,通过在琼脂板上对溶菌产物的连续稀释确定大量内化细菌(a,c)。如隔夜培养金黄色葡萄球菌6850(100cfu/ml)所示,通过施用u0126来分析u0126对细菌生长的影响。16小时后,将连续稀释物在bhi琼脂上(b,d)平板培养。细菌效价以线性(a,b)或对数刻度(c,d)描绘为菌落形成单位/ml(cfu/ml)。数据代表两种生物样品四个(a,c)或三个(b,d)独立实验的平均值±标准差(sd)。用双侧双样本t检验(***p<0.001)来评估统计学显著性。
图16:mek/erk信号传导的抑制引起病毒蛋白和促炎趋化因子的减少
在37℃,感染复数(moi=5)下,使人肺上皮细胞(a549)感染人流感病毒株a/威斯康星州/67/2005(h3n2)。30分钟后,将病毒稀释物移除,用pbs清洗细胞,并在50μmu0126或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌6850(moi=50)的侵染培养基dmem/inv(包含1%人血清白蛋白,25nmhepes)。细菌感染后3小时,用包含10%fbs,100μg/ml庆大霉素的dmem/fbs处理细胞30分钟以移除细胞外细菌。另外进行pbs清洗后,为细胞补充包含50μmu0126或溶剂的感染培养基dmem/ba(0.2%ba,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素)。37℃下培养8小时后,通过具有特异性引物的qrt-pcr来分析ccl3和ccl5的mrna水平(a,b)。通过蛋白质印迹分析确定病毒蛋白表达(pb-1)、erk-1/2磷酸化水平以及细菌细胞壁成分(pgn)(c)。数据代表一次实验中三个生物样本和两次技术重复的初步结果。
图17:u0126的施用引起独立于病毒效价的体内细菌效价降低
使12周大的balb/c小鼠在第0天感染50pfu流感病毒株a/波多黎各/8/34(pr8,h1n1)(用异氟烷麻醉)。从第一天开始每天对小鼠腹腔注射30mg/kg/dayu0126或溶剂对照(10%dmso,30%聚氧乙烯蓖麻油,60%pbs)。第三天,在异氟烷麻醉下使小鼠感染5*107cfu的金黄色酿脓葡萄球菌6850,并直接用u0126或溶剂对照处理。第四天,移除肺部并在pbs中均质化(0.1g/1000μlpbs)。将均质化物的连续稀释物在bhi琼脂上平板培养以计算细菌效价。进行标准空斑试验以确定病毒效价。用曼-惠特尼u检验进行统计学分析(*p<0.05)
图18:特定的mek抑制剂ci-1040降低单一和合并感染中的病毒效价
在37℃,感染复数(moi=0.01)下,使人肺上皮细胞(a549)感染禽流感病毒株a/fpv/布拉迪斯拉发/79(h7n7)(a,c)或人流感病毒a/波多黎各/8/34(h1n1)(b,d)。30分钟后,将病毒稀释物移除,用pbs清洗细胞,并在10μmci-1040或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌6850(moi=0.1)的侵染培养基dmem/inv(包含1%人血清白蛋白,25nmhepes)。细菌感染后3小时,用包含10%fbs,2μg/ml溶葡球菌酶的dmem/fbs处理细胞20分钟以移除细胞外细菌。另外进行pbs清洗后,为细胞补充包含10μmci-1040或溶剂的感染培养基dmem/ba(0.2%ba,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素)。37℃下培养18小时后,通过标准空斑试验确定病毒效价。病毒效价以线性(a,b)或对数刻度(c,d)描绘为血小板形成单位/ml(pfu/ml)。数据代表两种生物样品两个独立实验的平均值±标准差。
图19:ci-1040的施用对细菌的体外生长具有抑制效果
通过向隔夜培养的金黄色葡萄球菌6850(100cfu/ml)施用不同浓度的ci-1040(如所示的)来分析ci-1040对细菌生长的影响。16小时后,将连续稀释物在bhi琼脂上(a,b)平板培养。细菌效价以线性(a)或对数刻度(b)描绘为菌落形成单位/ml(cfu/ml)。数据代表两种生物样品的初步数据。
图20:用特定mek抑制剂考比替尼(gdc-0973)处理会降低体内病原体负荷
使8周大的balb/c小鼠(每组5只)在第0天感染50pfu的流感病毒株a/波多黎各/8/34(pr8,h1n1)(用异氟烷麻醉)。病毒感染6小时后,对小鼠进行口服施用10mg/kg/day考比替尼或溶剂对照进行治疗(10%dmso,5%吐温20,85%pbs)。然后每天进行治疗。第三天,在异氟烷麻醉下使小鼠感染5*107cfu的金黄色酿脓葡萄球菌6850,6小时后,用考比替尼或溶剂对照进行治疗。第四天,移除肺部并在pbs中均质化(0.1g/1000μlpbs)。将均质化物的连续稀释物在bhi琼脂上平板培养以计算细菌效价。进行标准空斑试验以确定病毒效价。用曼-惠特尼u检验进行统计学分析。
图21:lg-asa改善感染流感a病毒(iav)和/或金黄色酿脓葡萄球菌(金黄色葡萄球菌)的细胞形态
在37℃,感染复数(moi=0.1)下,使人肺上皮细胞(a549)溶解于pbs/ba[0.2%胎牛血清白蛋白,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素]感染流感病毒a/波多黎各/8/34(h1n1)。培养30分钟后,吸入病毒稀释物,用pbs清洗细胞,并在50mmlg-asa或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌sh1000(moi=0.1)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/lhepes]。细菌感染后3小时,用抗生素[dmem,10%fbs,2μg/ml溶葡球菌酶,20分钟]处理细胞以移除细胞外细菌。另外进行pbs清洗后,为细胞补充包含50mmlg-asa或溶剂的dmem/inv。37℃下培养8小时后,通过光学显微镜观察细胞形态。
图22:nfκb抑制剂lg-asa降低流感病毒效价和金黄色葡萄球菌负荷
在37℃,感染复数(moi=0.1)下,使人肺上皮细胞(a549)溶解于pbs/ba[0.2%胎牛血清白蛋白,1mmmgcl2,0.9mmcacl2,100u/ml盘尼西林,0.1mg/ml链霉素]感染流感病毒a/波多黎各/8/34(h1n1)(a-h)。培养30分钟后,吸入所述病毒稀释物,用pbs清洗细胞,并在5mmlg-asa或溶剂对照存在下为细胞补充含有或不含金黄色葡萄球菌sh1000(moi=50)(a,b),(moi=0.01)(c-d),金黄色葡萄球菌6850(moi=50)(e,f),(moi=0.1)(g,f)的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/ihepes]。细菌感染后3小时,用抗生素[dmem,10%fbs,2μg/ml溶葡球菌酶,20分钟]处理细胞以移除细胞外细菌。另外进行pbs清洗后,为细胞补充包含5mmlg-asa或溶剂的dmem/inv。37℃下培养8小时(a,b,e,f)或18小时(c,d,g,h)后,通过标准噬斑试验来测定iav效价(a,c,e,g)。通过低渗休克溶解细胞,并通过在琼脂板上连续稀释来确定内化细菌的数量(b,d,f,h)。数据代表两种生物样本三组(a-h)平均值±标准差。用双侧单样本t检验(*p<0.05;**p<0.01)来评估统计学显著性。
图23:nfκb抑制剂lg-asa降低流感病毒效价和金黄色葡萄球菌负荷
本图以不同的方式显示图22所得到的数据。特别地,任意设定每个实验未经处理的对照组为100%,然后计算平均值。用双侧单样本t检验(*p<0.05;**p<0.01)来评估统计学显著性。
图24:nfκb信号传导的抑制引起细菌内化降低
用5mm(a-d)和10mm(c,d)lg-asa预培养人肺上皮细胞(a549)4小时,然后在含有或不含溶解于dmem/inv[1%人血清蛋白,25nmol/lhepes]的指定数量的lg-asa下,使其感染金黄色葡萄球菌6850(moi=50)30-120分钟(a,b),感染usa300(moi=5)(c,d)120分钟。感染30-120分钟后,引入抗生素清洗[dmem,10%fbs,2μg/ml溶葡球菌酶,20分钟]以移除非内化细菌。用pbs清洗细胞三次并通过低渗休克溶解细胞。通过在琼脂板上连续稀释来确定内化细菌的数量(a-d)。数据(a,c)代表两种生物样本三组独立实验的平均值±标准差。在图24b,d中,任意设定每个实验所述未经处理的对照组为100%,然后计算所述平均值。用双侧单样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。
图25:nfκb信号传导的刺激引起细菌内化的增强
用5mmlg-asa和2.5ng/mltnf-α预培养人肺上皮细胞(a549)4小时,然后在含有或不含溶解于dmem/inv[1%人血清蛋白,25nmol/lhepes]的指定数量的lg-asa和tnf-α下,使其感染(a)金黄色葡萄球菌6850或(b)金黄色葡萄球菌usa300(moi=5)120分钟(a)。感染后120分钟,引入抗生素清洗[dmem,10%fbs,2μg/ml溶葡球菌酶,20分钟]以移除非内化细菌。用pbs清洗细胞三次并通过低渗休克溶解细胞。感染后120分钟,通过在琼脂板上连续稀释来确定内化细菌的数量(a)。(a)数据代表两种生物样本四组独立实验的平均值±标准差。先通过单因素方差分析随后通过多项比较试验来评估统计学显著性(*p<0.05;**p<0.01;***p<0.001)。(b)数据代表三种生物样本三组独立实验的平均值±标准差,通过任意设定每个实验未经处理的对照组为100%,然后计算平均值。
图26:用lg-asa治疗iav/金黄色葡萄球菌合并感染小鼠引起存活率增强、体重损失降低
(a)使balb/c小鼠(每组4只)在第0天感染50pfu的流感病毒a/波多黎各/8/34。流感病毒感染后第六天,再使小鼠感染108cfu金黄色葡萄球菌6850。流感病毒感染后第七天,每天通过吸入剂用lg-asa(1m,10分钟)治疗合并感染小鼠一次。观察14天的存活率。
而(4/4)(黑线)未经处理的合并感染小鼠在合并感染后一天死亡,(2/4)lg-asa治疗的合并感染小鼠存活(灰线)。
(b)描绘两组独立实验。使9周大的balb/c小鼠(每组4只)在第0天上午感染50pfu的流感病毒a/波多黎各/8/34(用异氟烷麻醉)。病毒感染后6小时,给小鼠称重并在吸入室中用雾化水或1mlg-asa治疗小鼠10分钟。在第一天、第二天和第三天与第0天的同一时间进行该治疗。在第三天上午,在异氟烷麻醉下,使小鼠感染5*107cfu的金黄色酿脓葡萄球菌6850。在第四天最后一次给小鼠称重。用曼-惠特尼u检验进行统计学分析。
发明详述
以下描述包括可用于理解本发明的信息。并不承认本文所提供的的任何信息为现有技术或与请求保护的发明相关,或者任何明确或隐含引用的出版物为现有技术。
如上所述,本发明涉及用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂。
此外,本发明涉及用在用于预防和/或治疗细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂。
本文所用“mek抑制剂”也可以被指定为丝裂原活化蛋白激酶(mapk)激酶抑制剂。众所周知,在mapk路径中,mapk激酶激酶(mapkkk)激活mapk激酶(mapkk),其转而激活mapk,mapk将信号转导至例如转录因子或其他激酶或效应器/信号转换蛋白;例如,参见fremin和meloche的图1(fremin和meloche(2010),j.hematol.oncol.11;3:8)。本发明的mek抑制剂优选抑制主体,诸如本文所述的哺乳动物或鸟的mek1/2。然而,可能本发明的mek抑制剂不仅抑制mek,优选抑制mek1/2,而且还抑制其上游激酶(即mapkkk),进而发挥双重抑制作用。不受理论的束缚,plx-4032可以是这样一种双重抑制剂。因此,在优选的方面,本发明的mek抑制剂可以是一种双重抑制剂,进而抑制mek,优选抑制mek1/2及其对应的上游mapkkk。mek1/2是ras/raf路径中的mapkk,ras/raf充当mapkkk,erk1/2充当mapk。
mek抑制剂可以是小分子、大分子、肽、寡核苷酸等等。所述mek抑制剂可以是蛋白质或其片段或核酸分子。所述术语mek抑制剂还包括所述mek抑制剂的药学上可接受的盐。
本领域普通技术人员能够确定一种化合物是否是mek抑制剂。在一个实施方案中,所述mek抑制剂选自表1所列的化合物/抑制剂。
本发明的mek抑制剂优选地选自u0126、plx-4032、azd6244、azd8330、as-703026、gsk-1120212、rdea-119、ro-5126766、ro-4987655、ci-1040、pd-0325901、gdc-0973、tak-733、pd98059、pd184352arry-438162和pf-3644022,优选地为azd8330、gsk-1120212、u0126、gdc-0973、ci-1040、pd0325901、arry-438162、pf-3644022和azd6244,最优选地为u0126、ci-1040、gdc-0973(考比替尼)、azd8330、gsk-1120212,最优选为u0126、gdc-0973、ci-1040、azd8330和gsk-1120212。
这些抑制剂中的一些在下表中做了进一步描述。
表1:mek抑制剂
还优选地选自plx-4032、azd6244、azd8330、gdc-0973、rdea119、gsk1120212、ro51267766、ro4987655、tak-733和as703026。更优选地选自azd6244、azd8330、gsk1120212和plx-4032或者选自pd-0325901、azd-6244、azd-8330和rdea-119。这些mek抑制剂为本领域为公众所知,例如,如表1所述(fremin和meloche(2010),j.hematol.oncol.11;3:8)。
也能够从arthur和ley(2013)先天免疫中的丝裂原活化蛋白激酶(mitogen-activatedproteinkinasesininnateimmunity),naturereviewsimmunology13,679–692(2013)中获得这些抑制剂的更多信息。
的确,如所附实施例所示,本文所述的mek抑制剂u0126和ci-1040在合并感染以及单独细菌感染情况下表现出一定的效果。
本发明还提供了用在用于预防和/或治疗合并感染或细菌感染的方法中的p38抑制剂。“p38map激酶抑制剂”为本领域所知。所述术语“p38抑制剂”、“p38激酶抑制剂”和“p38map激酶抑制剂”在本文中可交换使用。在本发明的上下文中,p38map激酶抑制剂抑制p38map激酶。优选地,p38map激酶抑制剂抑制p38map激酶的一种亚型,优选地抑制p38map激酶的四种亚型(α、β、γ或δ)中的一种,其中优选地抑制α亚型,更优选地抑制p38map激酶的两种亚型的任意组合,更加优选地抑制p38map激酶的三种亚型的任意组合,最优选地抑制p38map激酶的全部亚型或p38map激酶的α、β、γ和δ亚型。在一些实施方案中,所述p38map激酶抑制剂抑制参与炎性疾病、自身免疫性疾病、破坏性骨疾病、增生性疾病、传染性疾病、病毒性疾病或神经退行性疾病的p38的亚型。据报道,所述p38map激酶的α亚型参与炎症、增殖、分化和细胞凋亡,然而,p38β、p38δ和p38γ的生物功能尚未被完全理解。因此,本文优选p38map激酶抑制剂抑制α亚型。
所述p38map激酶抑制剂可以是小分子、大分子、肽、寡核苷酸等等。所述p38map激酶可以是蛋白质或其片段或者核酸分子。术语p38抑制剂还包括p38抑制剂的药学上可接受的盐。
本领域普通技术人员能够确定一种化合物是否是p38抑制剂。
本领域内有很多p38抑制剂的例子。美国专利5,965,583、6,040,320、6,147,096、6,214,830、6,469,174、6,521,655公开了作为p38抑制剂的化合物。美国专利6,410,540、6,476,031和6,448,257也公开了作为p38抑制剂的化合物。类似地,美国专利6,410,540、6,479,507和6,509,361公开了被称为p38抑制剂的化合物。美国公开的专利申请20020198214和20020132843公开了被称为p38抑制剂的化合物。另外的p38map激酶抑制剂是birb796bs(1-(5-叔丁基-2-p-甲苯基-2h-吡唑-3-基)-3-[4-(2-吗啉-4-基-乙氧基)-萘-1-基]-脲);进一步的p38map激酶抑制剂参见branger(2002),j.immunol.168:4070-4077或us6,319,921。
其他的p38map激酶抑制剂是amg548(amgen)、birb796(boehringeringelheim)、vx702(vertex/kissei)、scio469、scio323(sciosinc.)、sb681323(glaxosmithkline)、ph-797804(pfizer)和org-48762-0(organonnv);例如,参见lee和dominguez在currmedchem.2005;12(25):2979-2994中以及dominguez在curropindrugdiscovdevel.2005jul;8(4):421-430中所述的。
根据本发明,所述抑制剂可以表现出对p38map激酶上游或下游或者直接对p38map激酶的调节作用,其中优选后一种作用方式。调节p38map激酶活性的抑制剂的例子包括可降低p38map激酶的转录和/或翻译、可减少或抑制p38map激酶的翻译后修饰和/或细胞运输,或者可缩短p38map激酶的半衰期的抑制剂。所述抑制剂还可以与p38map激酶可逆或不可逆结合,抑制其活化作用,使其酶活性失活,否则会干扰其与下游底物的相互作用。
p38map激酶的四种亚型具有高水平的序列同源性。p38map激酶的α和β亚型紧密相关,而γ和δ亚型更相异。考虑到高度的结构相似性,具有抑制p38map激酶的一种亚型的某种化合物通常能抑制所述map激酶的其他亚型也就不足为奇了。因此,在一些实施方案中,对p38map激酶的α亚型特异性的p38map激酶抑制剂具有至少三类结构特征,其理论上允许亚型特异性抑制。
能够通过多种方法确定候选的p38map激酶抑制剂的选择性结合。p38map激酶的各种亚型的基因为本领域所知。本领域普通技术人员能够容易地复制和表达激酶的各种亚型,将其纯化,并进行与候选化合物进行结合研究以确定亚型结合特征。对p38map激酶的α亚型进行了该系列实验并由美国专利6,617,324b1提供。
mihara(2008),br.j.pharmacol.154(1):153-164中描述了另一种酶选择性试验。在本文的一些实施方案中,p38map激酶抑制剂抑制p38map激酶四种亚型中的一种,更优选地,其抑制p38map激酶的两种亚型的任意组合,更优选地,其抑制p38map激酶的三种亚型的任意组合,例如p38-α(mapk14)、-β(mapk11)、-γ(mapk12或erk6)。可选择的,但是也是优选的,其抑制p38map激酶的全部四种亚型。
在一个实施方案中,p38抑制剂选自表2(图8)所列的抑制剂。在另一个实施方案中,p38抑制剂选自sb202190、ly2228820、cay10571、sb203580、tie2激酶抑制剂、2-(4-氯苯基)-4-(氟苯基)-5-吡啶-4-基-1,2-二氢吡唑基-3-酮、cgh2466、sb220025、抗生素llz1640-2、tak715、sb202190盐酸盐、skf86002、amg548、cmpd-1、eo1428、jx401、ml3403、rwj67657、sb202190、sb203580、sb203580盐酸盐、sb239063、scio469、sx011、tak715、pamapimod、losmapimod(gw856553)、dilmapimod(sb681323)、vx702、vx745、doramapimod(birb796)、bms-582949、arry-797、ph797804、scio-469,优选vx-702、sb202190、pamapimod、losmapimod(gw856553)、dilmapimod(sb681323)、doramapimod(birb796)、bms-582949、arry-797、ph797804和scio-469。
也能够从arthur和ley(2013)先天免疫中的丝裂原活化蛋白激酶(mitogen-activatedproteinkinasesininnateimmunity),naturereviewsimmunology13,679–692(2013)中获得这些抑制剂的更多信息。
除了mek抑制剂和p38抑制剂,本发明还涉及用在用于预防和/或治疗本发明的合并感染或细菌感染的方法中的nfκb(nfkb/nfkappab)抑制剂。本领域普通技术人员能够确定一种化合物是否是nfκb抑制剂。
nfκb(活化的b细胞的核因子κ-轻链增强子)是一种控制dna转录的蛋白质复合体。几乎在所有动物细胞类型中都可发现nf-κb,并且其参与细胞对诸如压力、细胞因子、自由基、紫外线照射、氧化的ldl和细菌或病毒抗原的刺激的反应。脊椎动物nfκb转录复合体可以是任意由子单位p50(nfκb1)、p52(nfκb2)、c-rel、rela(p65)和relb形成的各种同源或异源二聚体(gilmoretd.(2006)oncogene25:6680–6684)。这些复合体结合至称之为κb位点的dna调节位点,通常激活特定的靶基因表达。在大多数细胞类型中,nf-κb二聚物通过与任何几种iκb抑制剂蛋白(iκbα,β,ε,γ,p105和p100)联合而以非活化形式位于细胞质中。为对大量刺激物响应,iκb迅速被磷酸化、泛素化并被蛋白酶体降解。然后释放的nf-κb二聚物转位至其能够调节特定基因表达的细胞核。
iκb的磷酸化和降解对nfκb复合体的调节很重要,该nfκb复合体由包含两种激酶子单位ikkα和ikkβ及其有关的叫做nemo(akaikkγ)的支架式调节蛋白的所述iκb激酶(ikk)复合物调节的(gilmore和herscovich(2006),nf-κb信号:785和计数癌基因的抑制剂,25,6887–6899)。尤其是如本发明的实施例3中所示,nf-κb信号传导对细菌(比如金黄色葡萄球菌)向细胞的内化作用也很重要。
根据本发明,抑制剂可以表现出对nfκb的上游或下游或者直接对nfκb的调节作用,其中优选所述后一种作用模式。调节nfκb活性的抑制剂的例子包括可以降低nfκb转录和/或翻译,或可以缩短nfκb半衰期的抑制剂。所述抑制剂还可以可逆或不可逆地结合nfκb、抑制其活化作用、使其丧失活性,否则会干扰其与下游目标,诸如目标基因的相互作用。nfκb抑制剂还可以抑制蛋白激酶,诸如通过如ikk抑制来抑制ikba磷酸化的分子。具有这样活性的化合物为sc-893、bms-345541,其可以作为参照化合物。nfκb抑制剂还可以抑制蛋白磷酸酶或抑制蛋白酶体或泛素化。同样可作为参照化合物的这样的nfκb抑制剂的例子包括蛋白质酪氨酸磷酸酶抑制剂、硼酸盐、硼替佐米、npi-0052。可选择地,nfκb抑制剂可以阻止nfκb的核转位或其结合至dna。同样可作为参照化合物的这样的抑制剂的例子包括sn50、脱氢环氧甲基醌霉素和nfκb诱骗寡核苷酸(decoyodns)。可以从gupta等人(2010)(gupta等人,(2010)通过小分子作为治疗策略抑制nfκb的活化作用,biochimbiophysacta,1799(10-12):775-787)获取nfκb抑制剂的进一步信息。
nfκb抑制剂可以是小分子、大分子、肽、寡核苷酸等等。nfκb抑制剂可以是蛋白质或其片段或核酸分子。术语nfκb抑制剂还包括nfκb抑制剂的药学上可接受的盐。在一个实施方案中,nfκb抑制剂选自图9和图10中的表3和表4所列的抑制剂/分子。在另一个实施方案中,nfκb抑制剂选自图9中的表3所列的抑制剂/分子。在另一个实施方案中,nfκb抑制剂选自图10中的表4所列的抑制剂/分子。
在另一个实施方案中,nfκb抑制剂选自lasag、sc75741(及其衍生物)、mg132、tpca-1、pctc、imd0354、木犀草素、咖啡酸苯乙酯、豆蔻明、pf184、ikk16、sc514、醉茄素a、牛蒡甙元、bay11-7085、psi、pr39、ro106-9920、bay11-7821、ml-130、南蛇藤醇、丹参酮iia、hu211、胶霉毒素、cid2858522、和厚朴酚、穿心莲内酯、10z-hymenialdisine、achp、扁塑藤素、柳氮磺胺吡啶、ml120b二盐酸盐、氨来占诺、9-甲基链霉戊二酰亚胺、n-硬脂酰植物鞘氨醇、2-(1,8-萘-2-基)-苯酚、5-氨基水杨酸、bay11-7085、3,4-二羟基肉桂酸乙酯、helanalin、nf-κb活化抑制剂ii、jsh-23、糖皮质激素受体调节剂、cpda、ppm-18、阿司匹林(asa)、吡咯烷二硫代羧酸铵盐、(r)-mg132、sc75741(及其衍生物)、楝酰胺、水杨酸钠、qnz、ps-1145、cay10512、硼替佐米、双水杨酯、白藜芦醇、lasag、脱氧精胍菌素、苏灵大、萨力多胺、agro-100、chs828和/或姜黄素,优选为硼替佐米、姜黄素、双水杨酯、白藜芦醇、水杨酸钠、lasag、asa、脱氧精胍菌素、苏灵大、萨力多胺、agro-100、chs828,更优选为sc75741(及其衍生物)、asa和lasag,最优选为lasag。
除sc75741外,术语“sc75741”或“sc75741(及其衍生物)”为sc75741的衍生物也被本发明涵盖。
通常,本领域技术人员知晓如何发现一种化合物是否是mek抑制剂、p38抑制剂和/或nfκb抑制剂。关于如何确定一种化合物是否是mek抑制剂和/或p38抑制剂的进一步的例子是分离所述mek和/或p38nfκb蛋白。所述蛋白可以从以下细胞中分离出来:mek和/或p38蛋白自然表达的细胞,或其已经通过寡核苷酸转染或以直接引导mek和/或p38蛋白表达的病毒感染过度表达的细胞。此外,mek和/或p38蛋白还可以重组表达。在分离所述蛋白时,本领域普通技术人员能够在存在或不存在潜在的mek和/或p38抑制剂的情况下测量激酶的活性。如果激酶在存在所述抑制剂下的活性低于不存在所述抑制剂下的活性,该抑制剂则分别为mek和/或p38。
如果直接作用在mek和/或p38上,抑制剂应表现出约5μm或更少,优选500nm或更少,更优选100nm或更少的ic50值。在相关的实施方案中,相对于p38-α亚型而言,所述抑制剂应表现出优选比用相同或可比较的试验对相同抑制剂对抗其他p38map激酶亚型测试所观察到的结果至少少10倍的ic50值。应当注意的是,ic50值为检测依赖性,根据测定的不同可以改变。考虑化合物的ic50值的相对关系比考虑其确切的值本身更加重要。
ic50是抑制酶的活性达到不存在抑制剂时所测量的活性的50%时的化合物的浓度。
使用与对照组相比引起50%降低的抑制剂的浓度计算ic50值。ic50值为检测依赖性,根据测定的不同可以改变。因此,ic50值是相对值。赋予特定抑制剂的值将有一般可比性并非基于绝对的基础。
将包含map和/或mapk激酶并用潜在的活化剂、抑制剂或调节剂处理的样品或试样与不含所述抑制剂、活化剂或调节剂的对照组样品进行比较以观察抑制程度。能够赋予对照组样品(未经抑制剂处理)100%的相对map和/或mapk激酶活性值。当所述map和/或mapk激酶活性值相对于所述对照组大约为80%,可选地为50%或25-0%时,会抑制map和/或mapk激酶。当所述map和/或mapk激酶活性值相对于所述对照组为110%,可选地为150%,可选地为200-500%或1000-3000%更高时,会活化map和/或mapk激酶的活化。本发明的示例性的map激酶结合活性试验为:map和/或mapk激酶配体污点试验(aymerich等人,investopthalmolvissci.42:3287-93,2001);map和/或mapk激酶亲和柱层析试验(alberdi等人,jbiolchem.274:31605-12,1999)和map和/或mapk激酶配体结合试验(alberdi等人,jbiolchem.274:31605-12,1999)。各自整体通过引用方式并入。
所述抑制剂的选择性还可以通过mihara(2008),br.j.pharmacol.154(1):153-164中所述的激酶选择性试验进行测量
关于nfκb抑制剂,人们能够测量例如未经处理的对照细胞中nfκb的靶基因的基因产品(蛋白质),并将这些靶基因产品的表达与已经以nfκb抑制剂处理的细胞进行比较。一些靶基因在oeckinghaus和ghosh(2009)的转录因子的nf-κb家族及其调节(thenf-κbfamilyoftranscriptionfactorsanditsregulation),coldspringharbperspectbiol.oct2009;1(4):a000034中有论述。
当用抑制剂处理细胞时,表达水平降低。其他的策略可以是通过免疫印迹试验检测ikba的降解以及p-p65积累和nfκb的核转位。还可以通过使用电泳迁移率变动试验(emsa)分析与经抑制剂处理的细胞相比,nfκb与未经处理的细胞的dna的相互作用。
还可以通过与参照化合物比较作用来分析分子的抑制性质。本文所提及的“参照化合物”意指可用作确定一种分子是否具有mek抑制剂、p38抑制剂和/或nfκb抑制剂性质的阳性对照。因此,本文所列的任何所述抑制剂也可以用作这样的参照化合物。可能的测试可以是这样的测试:用参照化合物处理例如受到刺激以活化所述mek、p38和或nfκb路径的细胞,并与感兴趣的化合物平行地放置在不同的孔中。
本发明所述的抑制剂可用在用于治疗和/或预防的方法中。因此,所述术语“治疗(treating)”或“治疗(treatment)”包括为了减轻或改善病症的目的向患有包含细菌感染和流感病毒感染的合并感染的患者优选地以药剂的形式施用mek抑制剂、p38抑制剂和/或nfκb抑制剂。类似地,包括为了减轻或改善病症的目的向患有细菌感染的患者优选地以药剂的形式施用mek抑制剂、p38抑制剂和/或nfκb抑制剂。
此外,本文所用术语“预防”是指以防止疾病为目的的任何医疗或公共健康程序。本文所用术语“预防(prevent)”、“预防(prevention)”和“预防(preventing)”是指降低获得或发展给定病症的风险,给定病症即包含流感病毒感染和细菌感染的合并感染或者单独的细菌感染。“预防”还意指降低或抑制患者的包含流感病毒感染和细菌感染的合并感染或者单独的细菌感染的再发生。
本发明所述抑制剂在治疗合并感染中有效。本文所用“合并感染”包含流感病毒感染和细菌感染。这样的合并感染可通过宿主(例如患者和/或单个细胞)同时感染细菌和流感病毒而发生。还可以是宿主(例如患者和/或单个细胞)同时感染一种或多种病毒颗粒和一种或多种细菌。然而,这样的合并感染还可以连续发生。在这种情形下,是宿主和/或细胞首先感染一种或多种病毒颗粒,然后所述相同的宿主和/或细胞感染一种或多种细菌,反之亦然。所述两种感染之间的时间段可以是最多14天、13天、12天、11天、10天、9天、8天、7天、6天、5天、4天、3天、2天、1天、12小时、6小时、3小时、1.5小时或最少30分钟。
这样的情况还可以是重复感染,其中二次感染叠加在较早的感染上,特别是通过对首次感染所用的治疗剂具有耐受性的外源或内源的不同菌剂。
在所述合并感染中,所述流感病毒感染可由流感a病毒或流感b病毒介导,优选地所述流感a病毒为h1n1、h2n2、h3n2、h6n1、h7n7、h7n9、h9n2h10n7、h10n8或h5n1。在一个实施方案中,所述流感a病毒为h1n1。在其他实施方案中,所述流感a病毒为h3n2、h5n1和h7n9。在另外的实施方案中,所述流感a病毒为h3n2、h5n1、h1n1和h7n9。
本发明还涉及“细菌感染”,其可在上述合并感染的环境下发生或可在仅有的感染存在于宿主(例如患者和/或细胞)中时发生。所述细菌感染可由任何细菌介导,优选地由以下细菌介导:葡萄球菌科、链球菌科、人菌科、假单胞菌科、衣原体科、支原体科、肠杆菌科、假单胞菌目和/或巴斯德氏菌科。
在其他实施方案中,所述细菌感染由以下细菌介导:葡萄球菌,优选为金黄色酿脓葡萄球菌;甲氧西林敏感性和抗甲氧西林耐受性金黄色葡萄球菌;杀白细胞素(pvl)-表达金黄色酿脓葡萄球菌和/或链球菌,优选为缓症链球菌、酿脓链球菌或肺炎链球菌;军团菌,优选为嗜肺军团菌;假单胞菌,优选为绿脓杆菌;衣原体,优选为肺炎衣原体;支原体,优选为肺炎支原体;克雷伯氏菌,优选为肺炎克雷伯氏菌;莫拉克斯氏菌,优选为卡他莫拉菌和/或嗜血杆菌,优选为流感嗜血杆菌。优选地,所述细菌选自金黄色酿脓葡萄球菌、肺炎链球菌或流感嗜血杆菌。最优选地,所述细菌为金黄色酿脓葡萄球菌。
本发明还涵盖抑制剂能够彼此联合。因此,在一个实施方案中,mek抑制剂与另一种mek抑制剂、p38抑制剂和/或nfκb抑制剂联合。在进一步的实施方案中,p38抑制剂与另一种p38抑制剂、mek抑制剂和/或nfκb抑制剂联合。在另一个实施方案中,nfκb抑制剂与另一种nfκb抑制剂、p38抑制剂和/或mek抑制剂联合。关于上述,术语“另一种抑制剂”用于阐明例如一种mek抑制剂还可与另外一种mek抑制剂联合,而这两种mek抑制剂是不同的。例如,mek抑制剂ci-1040可与mek抑制剂gdc-0973联合。这同样适用于p38和nfκb抑制剂。
在一个实施方案中,mek抑制剂、p38抑制剂和/或nfκb抑制剂与一种或多种针对流感病毒和细菌的另外的抑制剂同时施用、在前施用或在后施用。
在进一步的实施方案中,mek抑制剂,p38抑制剂和/或nfκb抑制剂用在用于预防和/或治疗本发明的合并感染的方法中,其中,所述mek抑制剂,p38抑制剂和/或nfκb抑制剂与针对所述流感病毒和/或所述细菌的一种或多种抑制剂联合。在一个实施方案中,所述mek抑制剂,p38抑制剂和/或nfκb抑制剂与针对所述流感病毒和/或所述细菌的一种或多种抑制剂同时施用、在前施用或在后施用。
通常,针对所述流感病毒的抑制剂是在流感治疗中有效的任何抑制剂或药剂。已知不同物质对降低流感感染有效。其中例如神经氨酸苷酶抑制剂、针对离子通道蛋白(m)的化合物和针对聚合酶或通过干扰所述病毒聚合酶复合体、pb1、pb2、pa或np组分的内切酶活动的化合物。本发明还涵盖了这些抑制剂药学上可接受的盐。
“神经氨酸苷酶抑制剂”是一种针对流感病毒的抗病毒药,其通过封锁病毒神经氨酸苷酶蛋白的功能,进而防止病毒结合到其意欲感染的细胞上和/或者防止病毒通过在宿主细胞发芽而繁殖来发挥作用,这是因为新生病毒不能在其进行复制的细胞中发芽成长。优选的神经氨酸苷酶抑制剂是奥司他韦、扎那米韦、帕拉米韦或这些物质中任何一个的药学上可接受的盐,比如奥司他韦磷酸盐、奥司他韦羧酸盐等。最优选的神经氨酸苷酶抑制剂为奥司他韦磷酸盐、扎那米韦、奥司他韦或帕拉米韦。
针对离子通道蛋白(m2)的化合物例如金刚烷胺和/或金刚烷乙胺,而针对聚合酶或通过干扰所述病毒聚合酶复合体、pb1、pb2、pa或np组分的内切酶活动的化合物例如np阻断剂nucleozin或聚合酶抑制剂t-705。
可选地或另外地,mek抑制剂,p38抑制剂和/或nfκb抑制剂可与一种或多种针对细菌的抑制剂联合。针对所述细菌的抑制剂可以是对降低细菌感染有效的任何抑制剂。本领域技术人员熟知的优选的抑制剂为抗生素。可以从表5(图11)获得优选的抗生素。因此,在一个实施方案中,所述抗生素选自表5(图11)所列的抗生素。在进一步的实施方案中,所述抗生素选自表5(图11)所列类别的抗生素。在另一个实施方案中,所述抗生素选自表5(图11)所列通用名的抗生素。更优选地,所述抗生素选自庆大霉素、利福平、lysosthaphin、红霉素、左氧氟沙星、万古霉素、替考拉宁、盘尼西林和苯甲异恶唑青霉素。
可经本发明所述抑制剂或抑制剂的组合治疗的“主体”优选为脊椎动物。在本发明上下文中,术语“主体”意指需要进行合并感染或单一细菌感染治疗的个体。优选地,所述主体是患有合并感染或单一细菌感染或存在感染风险的患者。优选地,所述患者为脊椎动物,更优选为哺乳动物。哺乳动物包括但不限于家畜、运动动物、宠物、灵长类、小鼠和大鼠。优选地,所述哺乳动物为人、狗、猫、母牛、猪、老鼠、大鼠等等,特别优选为人类。在一些实施方案中,所述主体为人类患者,其可选地为1岁以上14岁以下;50-65岁之间或者大于65岁。在其他实施方案中,所述主体为人类患者,其选自至少50岁的患者,处在长期护理设施下的患者,有肺或心血管慢性疾病的患者,在过去一年中因为慢性新陈代谢疾病、肾功能不全、血红蛋白病或免疫抑制需要定期医疗随访或住院治疗的患者,小于14岁的患者,正在接受长期阿司匹林治疗的6-18个月大的患者,以及在流感季将处于妊娠期第二个或第三个三个月期间的女性。
在本发明所述的方法中,所述mek抑制剂、p38抑制剂或nfκb抑制剂以及针对所述流感病毒的所述抑制剂和针对所述细菌的所述抑制剂可以口服施用、静脉注射、胸腔内给药、肌内注射、局部给药或吸入给药。优选地,所述mek抑制剂通过鼻吸入或口服施用。
本发明还涵盖不同的组合物。本发明涉及包含用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂的组合物。同样地,本发明涉及包含用在用于预防和/或治疗细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂的组合物。本发明还提供包含用在用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂以及针对所述流感病毒和/或所述细菌的一种或多种抑制剂的组合物。此外,本发明涉及包含用在用于预防和/或治疗细菌感染的方法中的mek抑制剂,p38抑制剂和/或nfκb抑制剂以及针对所述流感病毒和/或所述细菌的一种或多种抑制剂的组合物。
包含所述mek抑制剂,p38抑制剂和/或nfκb抑制剂以及另外的针对所述细菌的一种或多种抑制剂和/或针对所述流感病毒的一种或多种抑制剂的所述组合物可以是一种药物组合物。优选地,这样的组合物进一步包含载体,优选地为药学上可接受的载体。所述组合物可以是口服施用的混悬剂或片剂形式;鼻用喷雾剂,无菌注射剂(静脉注射、胸腔内给药、肌内注射),例如无菌注射水性或油性混悬剂或栓剂。在以混悬剂口服施用时,根据药剂领域可用的技术制备这些组合物并且可以包含用于扩容的微晶纤维素、作为助悬剂的海藻酸或海藻酸钠、作为黏度增强剂的甲基纤维素以及本领域所知的甜味剂/调味剂。作为直接释放的片剂,这些组合物可以包含微晶纤维素、磷酸氢钙、淀粉、硬脂酸镁和乳糖和/或其他本领域所知的赋形剂、粘结剂、填充剂、崩解剂、稀释剂和润滑剂。
所述可注射溶液剂或混悬剂可以根据已知技术使用适宜的无毒、不经肠道可接受的诸如甘露醇、1,3-丁二醇、水、格林氏溶液或等渗氯化钠溶液的稀释剂或溶剂,或者适宜的诸如包括合成的单甘脂或双甘酯的无菌温和不挥发油以及包括油酸的脂肪酸的分散剂或润湿剂或混悬剂进行制备。
所述抑制剂优选地以治疗有效量进行施用。用于本发明的并且包含mek抑制剂、p38抑制剂和/或nfkb抑制剂以及可选地一种或多种针对流感病毒的抑制剂和/或一种或多种针对细菌的抑制剂的药物组合物施用于哺乳动物或鸟类患者。合适的哺乳动物的实例,包括但不限于,小鼠、大鼠、奶牛、山羊、绵羊、猪、狗(dog)、猫、马、豚鼠、犬(canine)、仓鼠、貂、海豹、鲸、骆驼、黑猩猩、猕猴和人,优选为人。合适的鸟的例子,包括但不限于,火鸡、鸡、鹅、鸭子、水鸭、野鸭、八哥、北方针尾鸭、鸥、天鹅、珍珠鸡或水鸟,仅举几例。人类患者是本发明的具体的实施方式。
各活性化合物/抑制剂的“治疗有效量”随着但不限于以下因素而变化:所用化合物的活性、该活性化合物在患者体内的稳定性、待缓解的症状的严重程度、被治疗患者的总重量、给药途径、该活性化合物在体内吸收、分布、排泄的容易程度、被治疗患者的年龄和敏感性、不良反应等,这些对于本领域的技术人员而言都是显而易见的。给药量可以随时间依照这些因素进行调整。
本文所述抑制剂、方法及其用途既适用于人类治疗又适用于兽医应用。本文所述化合物,特别是具有理想的治疗活性的mek抑制剂、p38抑制剂和/或nfkb抑制剂以及可选地一种或多种针对流感病毒的抑制剂和/或一种或多种针对细菌的抑制剂可以如本文所述在生理上可接受的载体中向患者施用。根据引入方式,可以通过如下所述的多种方式制备所述化合物。制剂中治疗活性化合物的浓度可在约0.1-100wt%之间变化。试剂可单独施用或与其他治疗剂联合施用。
本发明方法中使用的药物化合物能够以任何合适的单位剂量形式给药。合适的口服剂型可以是片剂、胶囊剂、混悬液、糖浆剂、口香糖、薄片(wafer)、酏剂等。该口服药物组合物中可以包括药学上可接受的载体,如粘结剂、赋形剂、润滑剂、甜味剂和矫味剂。如果需要,还可以包括常规的改良特殊形式的味道、颜色及形状的制剂。
对于可注射类制剂,所述药物组合物可以是冻干粉混合有适当的赋形剂置于适宜的药瓶或管中。临床使用前,该药物可通过将冻干粉溶解在适宜的溶剂体系中进行重建,以形成适宜静脉注射或肌内注射的组合物。
根据本发明的另一个实施方案,本发明提供一种药物组合物,该药物组合物包含治疗有效量的mek抑制剂以及选自奥司他韦、奥司他韦磷酸盐、扎那米韦和帕拉米韦的治疗有效量的神经氨酸苷酶抑制剂。
在一个实施方案中,如前所述,所述组合物可以是可口服给药的形式(如片剂、胶囊剂或糖浆剂),包含治疗有效量(如0.1mg-2000mg、0.1mg-1000mg、0.1mg-500mg、0.1mg-200mg、30mg-300mg、0.1mg-75mg、0.1mg-30mg)的mek抑制剂、p38抑制剂和/或nfkb抑制剂以及治疗有效量(如0.1mg-2000mg、0.1mg-1000mg、0.1mg-500mg、0.1mg-200mg、30mg-300mg、0.1mg-75mg、0.1mg-30mg)的神经氨酸苷酶抑制剂。
在进一步的实施方案中,所述mek抑制剂、p38抑制剂和/或nfκb抑制剂用在用于预防和/或治疗本发明所述合并感染的方法中,其中,与接触前的体外测试系统相比,所述mek抑制剂、p38抑制剂和/或nfκb抑制剂在与体外测试系统接触时降低所述病毒和细菌感染,其中所述试验系统包含感染
a)流感病毒,和
b)细菌
的培养细胞。在另一个实施方案中,所述mek抑制剂、p38抑制剂和/或nfκb抑制剂用在用于预防和/或治疗本发明所述细菌感染的方法中,其中,与接触前的所述体外测试系统相比,所述mek抑制剂、p38抑制剂和/或nfκb抑制剂在与体外测试系统接触时降低所述细菌感染,其中所述试验系统包含感染细菌的培养细胞。
因此,本发明还涉及一种体外测试系统,其中所述试验系统包含感染
a)流感病毒,和
b)细菌
的培养细胞。沿着这样的思路,本发明还提供一种体外测试系统,其中所述试验系统包含感染细菌的培养细胞。
在所述体外测试系统包括病毒感染和细菌感染的情况下,同样,这些感染可先后发生或同时发生。
“培养细胞”(culturedcell)或“培养细胞“(culturedcells)是不存在于其自然环境(例如在植物或动物体内)中的细胞。培养细胞可以是包含与其自然环境分离的细胞或细胞系的原代细胞培养。优选地,所述培养细胞为人肺上皮细胞。优选地,所述培养细胞以0.5ml、1ml、1.5ml、2ml、2.5ml、3ml、3.5ml、4ml诸如dmem的培养基中约1x105、2x105、3x105、4x105、5x105、6x105、7x105、8x105、9x105、10x105、11x105、优选地8x105个细胞的密度接种。最优选地以2mldmem中8x105个细胞的密度接种。
使这样的培养细胞感染病毒和细菌或者在其他实施方案中使其只感染细菌。如上所述,合并感染可以先后或同时发生。例如,可以首先使所述培养细胞感染所述流感病毒,30分钟后使其感染细菌。3小时后,还可以另外向培养基中加入抗生素以移除细胞外细菌。在这样的情况下,要对所述抗生素再次进行清洗。在其他实施方案中,使所述细胞只感染细菌。
本文所述术语“接触”是指使包含流感病毒和细菌的细胞与mek抑制剂、p38抑制剂和/或nfκb抑制剂在空间上极为贴近。例如,这可以意指经由注射器将抑制剂应用于培养细胞处于其中的培养基。
然后,一旦接触,如果所述抑制剂具有活性,那么所述病毒感染以及所述细菌感染都会被降低。在一些实施方案中,在不存在病毒感染时本发明所述抑制剂用于降低细菌感染。
在一个实施方案中,所述病毒感染的降低是指血小板形成单位(pfu)/ml的降低,所述细菌感染的降低是指菌落形成单位(cfu)/ml的降低。所述血小板形成单位(pfu)/ml是指测量的每单位体积的能够形成血小板的颗粒(诸如病毒颗粒)的数量。它是一种功能性的测量而非颗粒绝对数量的测量:有缺陷的或未能感染其靶细胞的病毒颗粒不会产生血小板因而也不会计数。例如,浓度为1000pfu/μl的流感病毒溶液表明1μl的所述溶液包含足以在细胞单分子层产生1000个传染性血小板的病毒颗粒。在本发明的情况下,与用本发明所述的抑制剂处理前的培养基相比,用抑制剂处理过的细胞培养显示出经处理后培养基中血小板形成单位的数量降低。
通过以下方式分析可能的“血小板形成单位(pfu)/ml的降低”。首先,通过例如从培养皿吸入一些细胞并平板培养以计算形成的细菌血小板的数量来分析合并感染了流感病毒和细菌的培养细胞产生血小板形成单位(pfu)/ml的能力。然后将这一结果与应用了抑制剂后的相同培养基中的细胞产生的血小板形成单位(pfu)/ml的数量进行比较。如果与应用所述抑制剂之前产生的数量相比,在用抑制剂处理后所述血小板形成单位(pfu)/ml的数量降低,那么血小板形成单位(pfu)/ml降低。
“菌落形成单位(cfu)/ml”估算样本中的活菌数量。存在不同的方法。例如,为了产生菌落形成单位,将样本(例如,少体积的培养细胞)沿营养琼脂板的表面铺开并允许其在孵化前干燥以便计数。活菌被界定为在受控条件下经由二分裂进行繁殖的能力。当数菌落时不确定菌落是源自一个细胞或是1000个细胞时,细胞培养基中菌落的可见外观要求显著增长。因此,对于液体,结果报告为cfu/ml(每毫升菌落形成单位),对于固体,结果报告为cfu/g(每克菌落形成单位),从而反映这种不确定性(而不是细胞数/ml或细胞数/g)。
以下面的方式分析“菌落形成单位(cfu)/ml”。首先,通过例如从培养皿吸入一些细胞并平板培养以便计数来分析合并感染了流感病毒和细菌的所述培养细胞产生菌落形成单位(cfu)/ml的能力。然后将这一结果与应用所述抑制剂后的相同培养基中的细胞产生的菌落形成单位(cfu)/ml的数量进行比较。如果与应用所述抑制剂之前相比,在用抑制剂处理后所述菌落形成单位(cfu)/ml的数量降低,那么存在降低。
通常,本领域的技术人员知晓这些分析细菌和病毒感染的周知技术。文献中进一步描述了怎样能够测量所述血小板形成单位(pfu)/ml和所述菌落形成单位(cfu)/ml(tuchscherr,l.等人(2011),金黄色葡萄球菌菌相转换:一种逃离宿主免疫反应并建立慢性感染的有效的细菌策略,embo分子医学3,129-141和hrincius,e.r等人(2010),crk衔接蛋白表达对于禽流感a病毒的有效复制是必需的并且会控制jnk介导的凋亡反应,细胞微生物学12,831-843)。
此外,本发明涉及以下项目:
第1项、本发明还提供所述的体外测试系统用于确定抑制剂有效降低包含细菌感染和流感病毒感染的合并感染的用途。在一个实施方案中,所述病毒感染的降低是指血小板形成单位(pfu)/ml的降低,所述细菌感染的降低是指菌落形成单位(cfu)/ml的降低。
第2项、此外,本发明涉及一种检测有效预防和/或治疗包含细菌感染和流感病毒感染的合并感染的分子的方法,包含将感兴趣的化合物与本发明所述的体外测试系统接触,其中,与接触前的体外测试系统相比,所述感兴趣的化合物同时降低所述病毒和细菌感染。在一个实施方案中,所述病毒感染的降低是指血小板形成单位(pfu)/ml的降低,所述细菌感染的降低是指菌落形成单位(cfu)/ml的降低。
第3项、此外,本发明涉及所述体外测试系统用于确定抑制剂有效降低细菌感染的用途。
第4项、另外,本发明涉及所述体外测试系统用于检查先天宿主细胞反应的用途,其任选地包括检查信号传导水平、产生的细胞因子和趋化因子表达、细胞凋亡和坏死的诱导和/或调解健康和疾病的氧化还原止血作用。
第5项、本发明还提供一种检测有效预防和/或治疗细菌感染的分子的方法,包含将感兴趣的化合物与本发明所述的体外测试系统接触,其中,与接触前的体外测试系统相比,所述感兴趣的化合物降低了所述细菌感染。
第6项、本发明进一步涉及感染流感病毒和细菌的培养细胞。
第7项、还提供了感染细菌的培养细胞。
第8项、本发明还涉及一种用于预防和/或治疗主体中包含细菌感染和流感病毒感染的合并感染的方法,包含向所述主体施用治疗有效量的本发明所述的mek抑制剂、p38抑制剂和/或nfκb抑制剂或本发明所述的药物组合物。
第9项、本发明还提供所述的mek抑制剂、p38抑制剂和/或nfκb抑制剂或所述药物组合物用于制备药剂的用途。
第10项、此外,本发明涉及所述的mek抑制剂、p38抑制剂和/或nfκb抑制剂或所述药物组合物用于预防和/或治疗包含细菌感染和流感病毒感染的合并感染的用途。
第11项、类似地,本发明还提供一种用于预防和/或治疗主体中细菌感染的方法,包含向所述主体施用治疗有效量的本发明所述的mek抑制剂、p38抑制剂和/或nfκb抑制剂或本发明所述的药物组合物。
第12项、此外,本发明涉及所述的mek抑制剂、p38抑制剂和/或nfκb抑制剂或所述药物组合物用于预防和/或治疗细菌感染的用途。
***
须注意,如本文所用的单数形式“一种”、“一个”和“所述/该”包括复数指代,除非文中另有明确声明。因此,例如,提及“一种试剂”包括一种或多种这样的不同试剂,而提及“该方法”包括指代本领域普通技术人员已知的可以被修改或替代本文所述方法的等同步骤和方法。
本文所引用的所有出版物和专利都将通过整体引用并入本文。当通过引用并入的材料与本说明书存在矛盾或不一致时,本说明书将优先于任何此类材料
除非另有声明,否则一系列元素之前的术语“至少”理解为指代该系列中的每个元素。本领域技术人员仅仅使用常规实验就将得知或能够确定本文所述本发明的具体实施方式的许多等同物。这样的等同物意在被本发明涵盖。
纵观本发明说明书及接下来的权利要求书,除非语境需要,词语“包括(comprise)”及其变形“包括(comprises)”、“包括(comprising)”都将被理解为意指内含所述整体或步骤或整体或步骤的组,但是不排除任何其他整体或步骤或整体或步骤的组。本文所使用的术语“包括(comprising)”可以用“包含(containing)”替换,或者有时可以用“具有(having)”替换。
本文所使用的“由…组成(consistingof)”排除了未在权利要求部分(claimelement)中指定的任何元素、步骤或成分。本文所使用的“基本上由…组成(consistingessentiallyof)”未排除对权利要求的基本特征和新颖性特征没有实质影响的材料或步骤。在本文的各实施方式中,“包括(comprising)”、“由…组成(consistingof)”、“基本上由…组成(consistingessentiallyof)”中的任何一个可用另外两个术语中的一个替换。
贯穿本说明书的文本引用了若干文件。本文中所引用的每个文件(包括所有的专利、专利申请、科学出版物、生产厂家的说明书、使用指南等),无论上文还是下文,都特此通过引用整体并入本文。本文中的任何表述都不能解释为承认本发明无权优先于在先发明公开的内容。
具体实施方式
以下实施例进一步说明了本发明。然而这些实施例不能解读为限制本发明的范围。所述实施例限于说明性目的,本发明的范围仅由权利要求书限定。
实施例1
在过去几年中,对除了接种疫苗或用常规的抗病毒药物对抗iv(神经氨酸苷酶和m2阻断剂)以及用常规的抗生素对抗金黄色葡萄球菌以外的额外的和替代的治疗策略的需求稳步增加。同时,一些用于抗病毒干预的细胞因素已被认定为潜在靶点。完全相反,关于在细菌感染期间细胞因子的路径,以及其特别是通过降低细菌数量和/或启动加快细胞因子表达用于抗菌治疗的靶点的知识很少被理解,甚至更多是在iv合并感染出现的情况下。
我们建立了一种感染方案,其允许确定在iv和金黄色葡萄球菌合并感染时存在或不存在潜在抗病毒药物下:(1)子代病毒效价和(2)细胞内细菌效价以及(3)宿主防御机制的变化。在最初的方法中,我们研究了与溶剂对照相比较,在单一感染或合并感染情况下,mek抑制剂(u0126=50μm)、p38抑制剂(sb202190=10μm)和nfκb抑制剂(lasag=5mm)对抗iv和金黄色葡萄球菌的作用。作为对照,用病毒神经氨酸苷酶抑制剂奥司他韦(达菲)(2μm)与hepes相比较。对于病毒感染,我们使用人流感病毒a/波多黎各(puertorico)/8134(h1n1)或禽流感病毒a/fpv/布拉迪斯拉发(bratislava)/79(h7n7),而对于细菌感染,我们使用金黄色葡萄球菌菌株6850。感染过程(图1):
在6孔培养板(8x105个细胞/孔)中,将人肺上皮细胞接种于2mldmem[10%fcs]中。接种16-20小时后,冲洗细胞,并在37℃、指定的感染复数下用pbs/ba[0.2%胎牛血清白蛋白(bsa)、1mmmgcl2、0.9mmcacl2、100u/ml盘尼西林、0.1mg/ml链霉素](每6个孔500μl)或包含所述病毒的pbs/ba进行培养。培养30分钟后,吸入所述病毒稀释物,用pbs清洗细胞,并在存在或不存在所述测试化合物情况下在所指定的moi下为细胞补充含有或不含细菌的侵染培养基dmem/inv[1%人血清白蛋白,25nmol/lhepes](每6孔2ml)。细菌感染3小时后,用抗生素处理细胞以移除细胞外细菌。因此,用pbs清洗细胞并随后在37℃下用mem/inv抗生素[2μg/ml溶葡球菌酶(sigma)](1ml每6孔)培养20分钟。用pbs另外再清洗之后,为细胞补充包含所述测试物质的dmem/inv,并在37℃下培养指定次数。a/波多黎各(puertorico)/8/34dmem/inv再另外补充0.333μg/ml胰蛋白酶(英杰公司)。如(hrincius等人,2010,tuchscherr等人,2011)所述测定iv效价和细胞内细菌。
iv效价描绘为血小板形成单位(pfu)/ml,金黄色葡萄球菌效价描绘为菌落形成单位(cfu)/ml。数据代表两种生物样品二至三个独立实验的平均值±标准差。用双侧双样本t检验(*p<0.05;**p<0.01;***p<0.001)来评估统计学显著性。
在合并感染情况下,由于先天性免疫反应以及自噬和细胞调亡机制的变化,金黄色葡萄球菌的存在会影响iv复制,而iv的存在会影响所述金黄色葡萄球菌的细胞内数量。然而,正如所料,我们观察到u0126(图2、3)、sb202190(图4)和lg-asa(图5)对iv复制的抑制作用。有趣的是,在存在金黄色葡萄球菌下经这些抑制剂处理,也降低了病毒效价。而且令我们惊喜的是,与存在或不存在iv无关,在存在u0126、sb202190或lg-asa时,金黄色葡萄球菌的细胞内数量降低。另外一个令人惊讶的观察与u0126(50μm)存在下的细菌复制有关。在37℃下,于不含活细胞的dmem/inv中培养金黄色葡萄球菌时,细菌效价已经降低,但降低到与细胞感染期间相同的程度,这表明细菌对细胞因素的依赖(图2e)。
作为对照,我们研究了应用所述病毒神经氨酸苷酶抑制剂奥司他韦(达菲)(图6)时的病毒效价和细胞内细菌数量。尽管在缺少或存在金黄色葡萄球菌情况下,病毒效价显著降低,但是细胞内细菌数量增加。
该结果表明针对细胞因子的物质而非对抗病原体本身的物质作为对抗iv和金黄色葡萄球菌的合并感染的抗感染药物的巨大潜能。
实施例2
在进一步的实验中,研究了在单一感染或合并感染情况下与溶剂对照相比较,mek抑制剂u0126、ci-1040和考比替尼(gdc-0973)对抗流感a病毒(iav)和金黄色葡萄球菌6850感染的作用。
合并感染过程描绘为图12。为检验我们的实验设置中人肺上皮细胞的生存能力,用光学显微镜观察感染18小时的细胞形态(图13),这时确定病原体负荷(图14)。
如图13所示,以金黄色葡萄球菌6850(6850)、流感病毒株a/fpv/布拉迪斯拉发(bratislava)/79(h7n7)(fpv)或a/威斯康星州(wisconsin)/67/2005(h3n2)单一感染以及合并感染会引起轻微但可清晰检测的细胞损伤。在u0126(50μm)存在下,细胞层出现更少的损伤。
在人肺上皮细胞中确定单一感染或合并感染情况下,与溶剂对照相比较,mek抑制剂u0126(50μm)对抗流感病毒复制a/fpv/布拉迪斯拉发(bratislava)/79(h7n7)(fpv)或a/威斯康星州(wisconsin)/67/2005(h3n2)的作用(图14)。
mek/erk信号传导的抑制导致单一感染情况下经iav亚型h7n7和h3n2感染的病毒效价显著降低(图14)。在存在u0126的合并感染情况下,病毒效价在合并感染的情况下也降低,在h7n7/金黄色葡萄球菌合并感染的细胞中达到显著水平(图14)。
此外,分析了mek抑制剂u0126(50μm)对抗内化金黄色葡萄球菌6850的作用(图15a,c)。在本实验设置中,在存在抑制剂时细菌效价仅略有降低。
为进一步研究通常情况下u0126对细菌生长的作用,向金黄色葡萄球菌6850的无细胞隔夜培养基补充不同数量的u0126(10μm和50μm)或溶剂(图15b,d)。与溶剂对照相比,在u0126存在下细菌生长以浓度依赖的方式被抑制(图15b,d)。
由于促炎性细胞因子和趋化因子表达促成严重的炎症和组织损伤,在存在或不存在u0126(50μm)的感染实验中通过qrt-pcr分析各自的趋化因子,诸如ccl3,也叫巨噬细胞炎症蛋白1α(mip1α),以及ccl5,也叫rantes的mrna合成(图16a,b)。iav-诱导的ccl3mrna合成在存在金黄色葡萄球菌6850时增强而在存在u0126时降低。类似地,iav-诱导的ccl5mrna合成在存在金黄色葡萄球菌6850时降低,在存在u0126时进一步降低。
在蛋白质印迹分析中,通过使用特异性磷酸化erk1/2抗体来验证u0126对mek/erk信号传导的抑制作用(图16c)。此外,在存在mek/erk信号传导抑制情况下,观察到病毒蛋白合成(pb1)降低。
为在体内小鼠模型中验证u0126的抗病原体潜能。对感染流感病毒的小鼠不经处理或每天用u0126处理并以金黄色葡萄球菌6850重复感染(图17)。施用u0126导致体内细菌效价降低而与病毒效价无关。病毒效价没有降低可以解释为施用u0126较晚,当时病毒效价已经在感染过程中降低。前面的实验已经表明,在流感病毒感染前给予,抑制剂具有更高的抑制作用。然而,在应用u0126时,细菌效价显著降低。
在其他方法中,在人肺上皮细胞中测定单一感染或合并感染情况下,与溶剂对照相比,mek抑制剂ci-1040(10μm)对抗流感病毒复制a/fpv/布拉迪斯拉发(bratislava)/79(h7n7)(fpv)或a/波多黎各(puertorico)/8/34(h1n1)的作用(图18)。
通过ci-1040抑制mek/erk信号传导导致单一感染和合并感染的情况下一旦感染iav亚型h7n7和h1n1会降低病毒效价(图18)。
为进一步研究通常情况下ci-1040对细菌生长的作用,向金黄色葡萄球菌6850的无细胞隔夜培养基补充不同数量的ci-1040(1μm和10μm)或溶剂(图19a,b)。与溶剂对照相比,在存在ci-1040下细菌生长以浓度依赖的方式被轻微抑制(图19a,b)。
为验证另一种mek抑制剂的抗病原体潜能,在体内小鼠模型中测试考比替尼(cobimetinib),对感染流感病毒的小鼠不经处理或每天用考比替尼处理并以金黄色葡萄球菌6850重复感染(图20)。施用考比替尼导致体内病毒和细菌效价轻微但可清晰检测的降低。由于最近已经表明考比替尼的最大耐受剂量为30mg/kg/天。因此,通过使用比本实验所用的(10mg/kg/天)远低于最大耐受剂量的更高剂量可以改善抑制作用。
综上所述,结果表明不同mek抑制剂作为潜在的抗-iav/金黄色葡萄球菌物质。
实施例3
在进一步的实验中,研究了在单一感染或合并感染情况下nfκb-抑制剂lg-asa(lasag)对抗流感a病毒(iav)和金黄色葡萄球菌6850感染的作用。
合并感染描绘为图21(上半部分)。为检验人肺上皮细胞在缺少或存在lg-asa(5mm)下经感染的生存能力,用光学显微镜观察感染18小时的细胞形态(图21)。在缺少lg-asa的情况下(左面),以iav和/或金黄色葡萄球菌感染时会导致细胞损伤,而在存在lg-asa的情况下(右面),细胞形态得到改善。
在人肺上皮细胞中确定单一感染或合并感染情况下(图22/23)感染后8小时(图22/23a,b,e,f)和18小时(图22/23c,d,g,h),nfκb-抑制剂lg-asa(5mm)对抗流感病毒复制a/波多黎各(puertorico)/8/34(h1n1)的作用。两种不同的金黄色葡萄球菌株用于感染:(a)金黄色葡萄球菌sh1000(图22/23a-d)和(b)金黄色葡萄球菌6850(图22/23e-h)。
在存在lg-asa情况下,感染后8小时和18小时iav复制降低,而细菌效价的降低仅在感染后18小时可以看到。在图22中,结果以线性标度描绘。为使lg-asa的病原体抑制作用更加直观,将三组独立实验的未经处理的对照组人为设定为100%,并体现其平均值(图23)。
由于在细菌感染期间直接加入lg-asa,这些结果表明对细菌内化非常早的作用,其在持续释放和新的细菌内化过程中增强。
最近研究已经表明,nfκb是单核细胞金黄色葡萄球菌的吞噬作用所必需的(zhu等人,2014;exp.cellres.1;327(2):256-63)。基于这些发现以及我们自己的观察,我们想得知lg-asa是否会阻止细菌的摄取。我们在细菌感染前4小时对人肺上皮细胞进行预处理并在感染后高达2小时确定内化细菌。为确保只检测内化细菌,在细胞溶解前通过抗生素清洗移除非内化细菌。该数据表明金黄色葡萄球菌6850的时间依赖性摄取,其在存在lg-asa时被阻断(图24a,b)。进一步显示出lg-asa对细菌菌株金黄色葡萄球菌usa300的浓度依赖性抑制作用(图24c,d)。为使lg-asa的病原体抑制作用更加直观,这在图24a,c中可见,将三组独立实验的未经处理的对照组人为设定为100%,并体现其平均值(图24b,d)。
为验证nfκb介导的信号传导对金黄色葡萄球菌内化的重要性,在细菌感染前4小时,通过tnf-α刺激来诱导nfκb。nfκb的激活导致金黄色葡萄球菌6850和金黄色葡萄球菌usa300的摄取增强。作为对照,同时通过lg-asa阻断tnf-α诱发的激活,结果正如所料,其导致tnf-α所促进的细菌摄取得到抑制(图25)。
为验证体内小鼠模型中lg-asa的抗病原体潜能,对存在和缺少lg-asa情况下不同的合并感染设置进行测试。
如图26所见,用lg-asa处理iav/金黄色葡萄球菌合并感染的小鼠导致增强的存活率(图26a)并降低的体重损失(图26b)。
综上所述,我们的结果表明,诸如lg-asa的nfκb抑制剂在体外和体内作为抗-iav/金黄色葡萄球菌物质。
参考文献
borgeling,y.,schmolke,m.,viemann,d.,nordhoff,c.,roth,j.andludwig,s.(2014).inhibitionof38mitogen-activatedproteinkinaseimpairsinfluenzavirus-inducedprimaryandsecondaryhostgeneresponsesandprotectsmicefromlethalh5n1infection.thejournalofbiologicalchemistry289,13-27.
chertow,d.s.andmemoli,m.j.(2013).bacterialcoinfectionininfluenza:a·randroundsreview.jama:the'ournaloftheamericanmedicalassociation309,275-282.
droebner,k.,pleschka,s.,ludwig,s.andplanz,0.(2011).antiviralactivityofthemek-inhibitoru0126againstpandemich1nlvandhighlypathogenicavianinfluenzavirusinvitroandinvivo.antiviralresearch92,195-203.
dudek,s.e.,luig,c.,pauli,e.k.,schubert,u.andludwig,s.(2010).theclinicallyapprovedproteasomeinhibitorps-341efficientlyblocksinfluenzaavirusandvesicularstomatitisviruspropagationbyestablishinganantiviralstate.journalofvirology84,9439-9451,
ehrhardt,c.andludwig,s.(2009).anewplayerinadeadlygame:influenzavirusesandthepi3k/aktsignallingpathway.cellularmicrobiology11,863-871.
ehrhardt,c.,marjuki,h.,wolff,t.,nurnberg,b.,planz,0.,pleschka,s.andludwig,s.(2006).bivalentroleofthephosphatidylinositol-3-kinase(pi3k)duringinfluenzavirusinfectionandhostcelldefence.cellularmicrobiology8,1336-1348.
ehrhardt,c.,ruckle,a.,hrincius,e.r.,haasbach,e.,anhlan,d.,ahmann,k.,etal.(2013).thenf-kappabinhibitorsc75741efficientlyblocksinfluenzaviruspropagationandconfersahighbarrierfordevelopmentofviralresistance.cellularmicrobiology15,1198-1211.
ehrhardt,c.,wolff,t.andludwig,s.(2007a).activationofphosphatidylinositol3-kinasesignalingbythenonstructuralns1proteinisnotconservedamongtypeaandbinfluenzaviruses.journalofvirology81,12097-12100.
ehrhardt,c.,wolff,t.,pleschka,s.,planz,0.,beermann,w.,bode,j.g.,etal.(2007b).influenzaavirusns1proteinactivatesthepi3k/aktpathwaytomediateantiapoptoticsignalingresponses.journalofvirology81,3058-3067.
eierhoff,t.,hrincius,er.,rescher,u.,ludwig,s.andehrhardt,c.(2010).theepidermalgrowthfactorreceptor(egfr)promotesuptakeofinfluenzaaviruses(iav)intohostcells.plospathogens6,e1001099.
gillet,y.,vanhems,p.,lina,g.,bes,m.,vandenesch,f.,floret,d.andetienne,j.(2007).factorspredictingmortalityinnecrotizingcommunityacquiredpneumoniacausedbystaphylococcusaureuscontainingpanton-valentineleukocidin.clinicalinfectiousdiseases:anofficialpublicationoftheinfectiousdiseasessocietyofamerica45,315-321.
gong,j.,fang,h.,li,m.,liu,y.,yang,k.,liu,y.andxu,w.(2009).potentialtargetsandtheirrelevantinhibitorsinanti-influenzafields.currentmedicinalchemistry16,3716-3739.
grundmann,h.,aires-de-sousa,m.,boyce,j.andtiemersma,e.(2006).emergenceandresurgenceofmeticillin-resistantstaphylococcusaureusasapublic-healththreat.lancet368,874-885.
haasbach,e.,reiling,s.j.,ehrhardt,c.,droebner,k.,ruckle,a.,hrincius,e.r.,etal.(2013).thenf-kappabinhibitorsc75741protectsmiceagainsthighlypathogenicavianinfluenzaavirus.antiviralresearch99,336-344.
hayden,f.(2009).developingnewantiviralagentsforinfluenzatreatment:whatdoesthefuturehold?clinicalinfectiousdiseases:anofficialpublicationoftheinfectiousdiseasessocietyofamerica48suppl1,s3-13.
hayden,f.g.andhay,a.j.(1992).emergenceandtransmissionofinfluenzaavirusesresistanttoamantadineandrimantadine.currenttopicsinmicrobiologyandimmunology176,119-130.
iverson,a.r.,boyd,k.l.,mcauley,j.l.,piano,l.r.,hart,m.e.andmccullers,j.a.(2011).influenzavirusprimesmiceforpneumoniafromstaphylococcusaureus.thejournalofinfectiousdiseases203,880-888.′
lwao,y.,lshii,r.,tomita,y.,shibuya,y.,takano,t.,hung,w.c.,etal.(2012).theemergingst8methicillin-resistantstaphylococcusaureuscloneinthecommunityinjapan:associatedinfections,geneticdiversity,andcomparativegenomics.jinfectchemother18,228-240.
lee,lt.,lee,c.w.,tung,w.1-1.,wang,s.w.,lin,c.c.,shu,j.c.andyang,c.m.(2010).cooperationoftlr2withmyd88,pi3k,andradinlipoteichoicacid-inducedcpla2/cox-2-dependentairwayinflammatoryresponses.theamericanjournalofpathology176,1671-1684.
ludwig,s.(2009).targetingcellsignallingpathwaystofighttheflu:towardsaparadigmchangeinanti-influenzatherapy.thejournalofantimicrobialchemotherapy64,1-4.
ludwig,s.(2011).disruptionofvirus-hostcellinteractionsandcellsignalingpathwaysasananti-viralapproachagainstinfluenzavirusinfections.biologicalchemistry392,837-847.
ludwig,s.andplanz,0.(2008).influenzavirusesandthenf-kappabsignalingpathway-towardsanovelconceptofantiviraltherapy.biologicalchemistry389,1307-1312.
ludwig,s.,planz,0.,pleschka,s.andwolff,t.(2003).influenza-virusinducedsignalingcascades:targetsforantiviraltherapy?trendsinmolecularmedicine9,46-52.
ludwig,s.,wolff,t.,ehrhardt,c.,wurzer,w.j.,reinhardt,j.,planz,0.andpleschka,s.(2004).mekinhibitionimpairsinfluenzabviruspropagationwithoutemergenceofresistantvariants.febsletters561,37-43.
marjuki,h.,alam,m.i.,ehrhardt,c.,wagner,r.,planz,0.,klenk,h.d.,etal.(2006).membraneaccumulationofinfluenzaavirushemagglutinintriggersnuclearexportoftheviralgenomeviaproteinkinasecalphamediatedactivationoferksignaling.thejournalofbiologicalchemistry281,16707-16715.
mazur,1.,wurzer,w.j.,ehrhardt,c.,pleschka,s.,puthavathana,p.,silberzahn,t.,etal.(2007).acetylsalicylicacid(asa)blocksinfluenzaviruspropagationviaitsnf-kappab-inhibitingactivity.cellularmicrobiology9,1683-1694.
moran,g.j.,krishnadasan,a.,gorwitz,r.j.,fosheim,g.e.,mcdougal,l.k.,carey,r.b.,etal.(2006).methicillin-resistants.aureusinfectionsamongpatientsintheemergencydepartment.thenewenglandjournalofmedicine355,666-674.
morens,d.m.,taubenberger,j.k.andfauci,a.s.(2008).predominantroteofbacterialpneumoniaasacauseofdeathinpandemicinfluenza:implicationsforpandemicinfluenzapreparedness.thejournalofinfectiousdiseases198,962-970.
neumann,g.,noda,t.andkawaoka,y.(2009).emergenceandpandemicpotentialofswine-originh1n1influenzavirus.nature459,931-939.
niemann,s.,ehrhardt,c.,medina,e.,warnking,k.,tuchscherr,l.,heitmann,v.,etal.(2012).combinedactionofinfluenzavirusandstaphylococcusaureuspanton-valentineleukocidinprovokesseverelungepitheliumdamage.thejournalofinfectiousdiseases206,1138-1148.
olschlager,v.,pleschka,s.,fischer,t.,rziha,h.j.,wurzer,w.,stitz,l.,etal.(2004).lung-specificexpressionofactiverafkinaseresuitsinincreasedmortalityofinfluenzaavirus-infectedmice.oncogene23,6639-6646.
oviedo-boyso,j.,cortes-vieyra,r.,huante-mendoza,a.,yu,h.b.,valdez-alarcon,j.j.,bravo-patino,a.,etal.(2011).thephosphoinositide-3-kinase-aktsignalingpathwayisimportantforstaphylococcusaureusinternalizationbyendothelialcells.infectionandimmunity79,4569-4577.
paddock,c.d.,liu,l.,denison,a.m.,bartlett,j.h.,holman,r.c.,deleon-carnes,m.,etal.(2012).myocardialinjuryandbacterialpneumoniacontributetothepathogenesisoffatalinfluenzabvirusinfection.thejournalofinfectiousdiseases205,895-905.
park,b.andliu,g.y.(2012).targetingthehost-pathogeninterfacefortreatmentofstaphylococcusaureusinfection.seminarsinimmunopathology34,299-315.
parker,d.andprince,a.(2012).immunopathogenesisofstaphylococcusaureuspulmonaryinfection.seminarsinimmunopathology34,281-297.
parry,j.(2013).h7n9avianfluinfectshumansforthefirsttime.bmj346,f2151.
pinto,l.h.andlamb,r.a.(2006).them2protonchannelsofinfluenzaaandbviruses.thejournalofbiologicalchemistry281,8997-9000.
pinto,l.h.andlamb,r.a.(2007).controllinginfluenzavirusreplicationbyinhibitingitsprotonchannel.molecularbiosystems3,18-23.
planz,0.(2013).developmentofcellularsignalingpathwayinhibitorsasnewantiviralsagainstinfluenza.antiviralresearch98,457-468.
pleschka,s.,wolff,t.,ehrhardt,c.,hobom,g.,planz,0.,rapp,u.r.andludwig,s.(2001).influenzaviruspropagationisimpairedbyinhibitionoftheraf/mek/erksignallingcascade.naturecellbiology3,301-305.
shilo,n.andquach,c.(2011).pulmonaryinfectionsandcommunityassociatedmethicillinresistantstaphylococcusaureus:adangerousmix?paediatricrespiratoryreviews12,182-189.
taubenberger,j.k,andkash,j.c.(2010).influenzavirusevolution,hostadaptation,andpandemicformation.cellhostµbe7,440-451.
thorburn,k.andriordan,a.(2012).pulmonarybacterialcoinfectionininfantsandchildrenwithviralrespiratoryinfection.expertreviewofantiinfectivetherapy10,909-916.
wurzer,w.j.,ehrhardt,c.,pleschka,s.,berberich-siebelt,f.,wolff,t.,walczak,h.,etal.(2004).nf-kappab-dependentinductionoftumornecrosisfactor-relatedapoptosis-inducingligand(trail)andfas/fasliscrucialforefficientinfluenzaviruspropagation.thejournalofbiologicalchemistry279,30931-30937.
zhuf,yuew,wangy.thenuclearfactorkappab(nf-κb)activationisrequiredforphagocytosisofstaphylococcusaureusbyraw264.7cells.expcellres.2014oct1;327(2):256-63.
- 还没有人留言评论。精彩留言会获得点赞!
- 人源抗H7N9禽流感病毒中和性抗体1F7L及其应用的制造方法与工艺
- 吲哚衍生物及其制备方法和其抗流感病毒作用与制造工艺
- 检测H5亚型禽流感病毒与鸡细小病毒的成套试剂与方法与制造工艺
- 检测禽流感病毒与鸡细小病毒的成套试剂与方法与制造工艺
- 一种流感病毒逆转录环介导等温扩增试剂盒及其应用的制造方法与工艺
- 用于快速检测流感病毒的狗肾细胞MDCK-NS1的离心小瓶及其应用方法与制造工艺
- 一种保护和稳定病毒RNA的采血管的制造方法与工艺
- H6亚型禽流感病毒荧光定量RT‑PCR检测方法与制造工艺
- 表达H7N9亚型禽流感病毒血凝素蛋白的重组火鸡疱疹病毒株rHOH及构建方法与制造工艺
- 一种提高禽流感疫苗毒株增殖滴度的方法与制造工艺