一种甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因及应用的制作方法
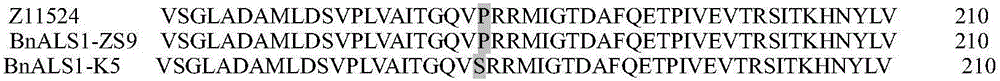
本发明涉及植物基因工程技术领域,具体涉及甘蓝型油菜抗磺酰脲类除草剂基因及其应用。
背景技术:
油菜是我国最重要的油料作物。农田杂草严重影响油菜产量和品质,是油菜生产主要生物灾害之一。抗除草剂品种及与其配套除草剂的结合使用是防除农田杂草最为经济有限的措施。北美、欧洲等发达国家早已种植抗除草剂油菜品种实施化学除草。目前,抗除草剂的油菜品种类型主要有孟山都公司注册的抗草甘膦油菜、拜耳公司的抗草铵膦油菜和巴斯夫公司的抗咪唑啉酮油菜。上述材料中的抗性基因均受国际知识产权保护,商业化种植需要交纳昂贵的专利费;而且抗草甘膦油菜和抗草铵膦油菜均为转基因品种。在我国油菜转基因品种尚未批准商业化种植,因此无法在生产上应用和推广。目前,我国缺乏商业化的抗除草剂油菜品种及其配套的除草剂,这已成为制约我国油菜产业发展的瓶颈。因此,创制具有自主知识产权、适合我国生产的抗除草剂油菜,研制配套的除草剂配方对我国油菜产业发展具有十分重要的意义。
磺酰脲(SU)类除草剂是目前世界上开发品种最多、应用范围最广的一类除草剂。其具有诸多优点:1)高效低量;SU类除草剂的除草效率极高,使用量极低,每公顷仅需几克到几十克,因而被称为超高效除草剂;2)杀草谱广;能有效防除双子叶阔叶杂草,其中有些除草剂品种对禾本科杂草也有很好的防效;3)土壤残留期短,基本不会影响后茬作物生长;4)环境友好,对哺乳动物毒性低,人及牧畜十分安全。然而,许多商业化的SU类除草剂对油菜具有强烈的生长抑制作用,致使其仅能在水稻、小麦、玉米等单子叶作物田间应用。目前,油菜田杂草防除采取的主要措施是播后苗前利用乙草胺等进行封闭除草。适于油菜田苗期防除单子叶杂草的除草剂品种比较齐全,而防除阔叶杂草的除草剂种类极少、且用药时期要求严格。选育高抗SU类除草剂的油菜品种,将SU类除草剂和现有油菜田单子叶除草剂混合使用,将拓宽现有SU类除草剂的应用范围,为有效防除油菜田杂草提供一条新的途径。然而,由于缺少生产应用价值的抗SU类除草剂油菜种质,国内至今还没有培育出商业化的抗SU类除草剂油菜品种。因此,筛选鉴定具有生产应用价值的油菜抗性材料(基因),创造高抗SU类除草剂油菜新种质,是我国实现油菜田间杂草化学防除的当务之急。
乙酰乳酸合成酶(ALS)及其抑制剂类除草剂
乙酰乳酸合成酶(ALS,acetolactate synthase),或称作乙酰羟酸合成酶(acetohydroxyacid synthase,AHAS),是催化植物和微生物体内支链氨基酸(包括亮氨酸、异亮氨酸、缬氨酸)生物合成途径的第一个关键限速酶。该酶同时催化2个平行反应:催化两分子丙酮酸缩合形成2-乙酰乳酸并释放出CO2,最终生成缬氨酸和亮氨酸;催化一分子丙酮酸和一分子2-氧丁酸缩合形成乙酰羟基丁酸,最终生成异亮氨酸。甘蓝型油菜中共有5个ALS基因。其中BnALS1和BnALS3氨基酸同源性达98%,属于看家基因,在根、茎、叶、花、角果中表达;BnALS2与BnALS1和BnALS3核苷酸同源性为85%,氨基酸同源性为75%,其仅在花、角果中表达;BnALS4和BnALS5是缺失的假基因。
以ALS为作用靶标的除草剂是一类非常重要的除草剂(Delye et al.Trends in Genetics,2013,29(11):649-658)。该类除草剂包括磺酰脲类(SU)、磺酰胺羰基三唑啉酮类(SCT)、咪唑啉酮类(IMI)、三唑嘧啶类(TP)和嘧啶水杨酸类(PTB)(Mallory-Simth and Retzinger,Weed Technol,2003,17:605-619)等五大类除草剂。其中SU类除草剂是目前世界上开发品种最多、应用范围最广的一类除草剂。目前在我国注册的SU类除草剂有近30个,其中,苯磺隆就是一个麦田防除阔叶杂草的代表性除草剂。这些SU类除草剂仅胺苯磺隆在油菜田注册使用,其它商业化的SU类除草剂对双子叶作物油菜具有强烈的生长抑制作用,致使其仅能在水稻、小麦、玉米等单子叶作物大田中应用。
抗除草剂植物的抗性机理
植物对SU类除草剂的抗性策略主要包括靶标抗性(Target-site resistance,TSR)及非靶标抗性(Non-target-site resistance,NTSR)(Delye et al.Trends in Genetics,2013,29(11):649-658)。
(1)靶标抗性(TSR):
靶标抗性是指通过靶标蛋白的表达活性升高或靶标蛋白与除草剂结合位点的结构改变而获得的抗性。除草剂靶标ALS氨基酸的替换是植物抗性产生的主要原因。不同位点氨基酸的替换以及氨基酸替换方式导致产生的抗性效应和交叉抗性均有较大差异。目前报道的ALS基因产生抗性突变的位点(以拟南芥ALS氨基酸序列作为参比)主要有以下8个:
Ala122、Pro197、Ala205、Asp376、Arg377、Trp574、Ser653和Gly654(http://www.weedscience.com,9 January 2014)。
上述这些位点的氨基酸种类在绝大多数已知植物的ALS序列中相当保守;其中,能引起植物对SU类除草剂产生抗性变化的氨基酸位点主要有3个,即Pro197、Ala205和Trp574。这3个位点氨基酸替换引起植物对SU类除草剂的抗性效应与交叉抗性存在显著差异。Pro197位点的氨基酸替换可导致植物对SU类除草剂具有高度抗性;且Pro-197-Leu变异也对IMI产生较高水平抗性。Ala205的变异对ALS抑制剂类除草剂均可以产生较高水平抗性。Trp574的氨基酸变异对IMI、SU和TP均产生很高的交叉抗性。此外,Ala122、Ser653和Gly654位点的氨基酸替换也致使植物对IMI产生高水平抗性;其中Ser-653-Thr的变异致使植物对SU也产生一定程度的抗性。由此可见,植物的抗性效应取决于ALS序列上氨基酸替换发生的位点以及替换氨基酸的种类。目前这方面研究多集中在田间抗性杂草上,作物中的相关研究甚少(Tranel et al.http://www.weedscience.com.2015.09.03)。
(2)非靶标抗性(NTSR):
非靶标基因的表达调控促进代谢作用,从而引起植物对除草剂产生抗性。非靶标抗性指除TSR以外的其他机制。研究表明至少4种酶系统,细胞色素P450酶系、谷胱甘肽转移酶(GSTs)、葡萄糖基转移酶和ABC转运蛋白参与了植物对SU类除草剂的代谢作用。水稻、小麦、玉米等禾本科作物大田能安全使用多数的SU类选择性除草剂,就是利用了这类酶系统对除草剂的快速代谢功能。在这几种酶系统中,P450酶系是将除草剂分解成无毒化合物的主要酶系。P450酶系相关基因的表达量增加,促进代谢除草剂能力提升,导致植物对SU类除草剂产生抗性。对于作用靶标单一而专化的SU类除草剂来说,这种由非靶标基因突变产生的抗性目前仅在陆地棉中有一例报道。
抗除草剂油菜
加拿大抗除草剂油菜育种及推广应用走在世界前列。目前抗除草剂油菜品种种植面积占油菜总面积95%以上。其中抗草铵膦品种占46%,抗草甘膦品种占47%,这二者均为转基因品种;非转基因的抗咪唑啉酮类除草剂品种占6%。澳大利亚主要种植非转基因的抗咪唑啉酮类除草剂的油菜。其它国家还未允许转基因油菜品种注册种植。
在非转基因抗除草剂作物品种选育及推广应用中,最为成功的就是以ALS基因为靶标,经过诱变和选择,开发得到的抗咪唑啉酮类除草剂作物。抗咪唑啉酮类作物及其配套的除草剂被称为Clearfield作物生产系统。目前,已选育成功玉米、油菜、水稻、小麦和向日葵的Clearfield系统,并登记上市。Swanson等(Plant Cell Rep,1988,7(2):83-87;Theor Appl Genet,1989,78(4):525-530)通过小孢子诱变筛选出两个抗咪唑啉酮类除草剂油菜突变体,分别命名为PM1和PM2;研究表明,这两个突变体都是由于ALS基因点突变产生的。其中PM1的抗性是由于BnALS1的Ser-653-Asp替换所致,PM2的抗性是由于BnALS3的Trp-574-Leu替换而产生;而且PM1只对IMI类除草剂有抗药性,而PM2同时对IMI类和SU类除草剂有抗性(Hattori et al.Mol Gen Genet,1995,246:419-425)。
巴斯夫公司以PM1和PM2为基础材料,1995年选育出抗IMI的Smart Canola,注册为Clearfield油菜。
我国江苏农科院浦惠明课题组于2004年在油菜和大豆多年轮作的试验田发现了自然突变的抗IMI的油菜材料M9(高建芹等,植物遗传资源学报,2010,11(3):369-373),其抗性性状为单基因控制的显性性状;是BnALS1蛋白的Ser-653-Asp替换所致(胡茂龙等,中国农业科学,2012,45(20):4326-4334;Hu et al.Mol Breeding,2015,35:1-13)。
浦惠明课题组还利用诱变剂甲基磺酸乙酯(EMS)处理甘蓝型油菜,获得了抗SU类除草剂突变体M343(中国专利申请号:201310054645.9),突变位点是BnALS3蛋白的Trp-574-Leu替换而产生(胡茂龙等,中国专利号:ZL201310111739.5)。
中国农科院油料所李小童等(中国油料作物学报,2015,37(3):269-276)报道的抗IMI的油菜种质12WH318也是BnALS1蛋白的Ser-653-Asp替换所致。
华中农业大学刘克德等用EMS处理甘蓝型油菜品种“华双5号”,筛选得到苯磺隆抗性材料,研究表明该性状受一对核基因控制,抗性产生是由于BnALS3基因第+535位C→T突变导致所编码BnALS3的Pro-197-Ser变异(Li et al.Theor Appl Genet,2015,128(1):107-118)。目前,油菜中已报道的ALS基因突变位点及特性详见表1。
表1:已报道的油菜ALS基因突变及其特性
a:表中突变位点均以拟南芥ALS氨基酸序列为参比;Trp(野生型)-574-Leu(突变型);ENU,ethyl nitrosourea;EMS,Ethylmethylsulfone.IMI,imidazolinones;SU,sulfonylureas,TP,triazolopyrimidines未报道。
技术实现要素:
为了弥补上述已有技术或专利的不足,本发明的目的在于,提供一种甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因及其应用。
为了实现上述任务,本发明通过以下技术方案得以实现:
一种甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因,其特征在于,该甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因由1968个碱基组成,序列为SEQIDNO.1,命名为Bnals1544T基因,即抗磺酰脲类除草剂苯磺隆基因,编码甘蓝型油菜乙酰乳酸合成酶I;与其它来源的油菜乙酰乳酸合成酶I的核酸序列相比,Bnals1544T基因包含一处突变:即C544T。
上述甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因编码的蛋白质由655 个氨基酸残基组成,序列为SEQIDNO.2,编码蛋白质序列第182或等同位置处的氨基酸残基为丝氨酸。
经申请人的研究表明,上述甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因可以用于培育抗除草剂植物的应用。
将甘蓝型油菜抗磺酰脲类除草剂苯磺隆Bnals1544T基因导入植物细胞或组织(拟南芥或油菜),获得对除草剂具备抗性的转基因植物。
本发明的甘蓝型油菜抗苯磺隆基因Bnals1544T的应用研究包括但不限于,应用植物常规育种方法将该基因导入其它对除草剂苯磺隆无抗性的目标植物,提高导入目标植物对除草剂苯磺隆的耐受性。根据本发明的甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因序列特征,设计分子标记,检测甘蓝型油菜抗性基因Bnals1544T的存在。
本发明的创新点在于:
1、甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因Bnals1544T是甘蓝型油菜中首次发现的。
2、利用本发明的甘蓝型油菜抗苯磺隆基因Bnals1544T的核酸序列,可以提高其它对苯磺隆无抗性的植物对苯磺隆的耐受性。当Bnals1544T基因在植物中表达时,其赋予植物对除草剂苯磺隆的抗性,由于除草剂苯磺隆土壤残留期短,基本不会影响后茬作物生长。因此,利用甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因Bnals1544T选育的油菜新品种,均能在我国油菜生产上推广应用。
附图说明
图1是甘蓝型油菜K5突变体的抗性特征。
图2是甘蓝型油菜K5的突变位点。
图3是甘蓝型油菜K5突变体Bnals1544T转基因拟南芥的PCR鉴定。其中M为DM2000Plus marker(CWBIO,Cat:cw2193),1-15为转基因拟南芥喷施草铵膦筛选后植株的PCR鉴定结果,其中有3株(NO.8,11,12)为假阳性植株,其余12株为转基因成功的阳性植株。
图4是抗性基因Bnals1544T的转拟南芥验证。图为拟南芥喷施2μg/ml苯磺隆水溶液结果,其中A、B、C为野生型拟南芥,a、b、c为阳性转基因植株。
以下结合附图和实施例对本发明做进一步的详细说明。
具体实施方式
申请人的研究小组利用EMS处理甘蓝型油菜品种“中双9号”构建了一个油菜突变体库;并通过苯磺隆抗性筛选在约30000株M2群体中筛选到3株苯磺隆抗性突变体K1、K4和K5,经申请人进一步研究发现,K5是BnALS1基因第+544位C→T突变(该突变基因简写为Bnals1544T)导致所编码产物BnALS1蛋白的Pro-182-Ser改变。K5纯合体抗苯磺隆安全浓度为100mg ai/L,最高耐受浓度为200mg ai/L。据申请人所知,油菜K5纯合体中的突变基因Bnals1544T是以前未报道的、申请人首次获得的一个抗除草剂候选基因。
以下发明人以获得的甘蓝型油菜抗苯磺隆除草剂突变体K5为材料,提供甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因核酸序列的分离、载体构建,以及培育具有对磺酰脲类除草剂苯磺隆抗性植物的具体实例。
需要说明的是,下列实施中所有方法如无特别说明均为常规方法。
(一)甘蓝型油菜抗除草剂苯磺隆基因Bnals1544T的获得
甘蓝型油菜K5突变体在田间苯磺隆除草浓度作用下,仍可正常生长,且可维持完整的发育进程(图1)。对照组(CK)为中双9号甘蓝型油菜喷施5μg/ml苯磺隆水溶液后,表现出叶面枯黄、叶裂加深、生长逐渐停止,直至植物枯萎死亡;实验组K5突变体喷施浓度为200μg/ml的苯磺隆水溶液后,仍然表现正常的生长状态,证明K5突变体对苯磺隆具有较高抗性。
对苯磺隆抗性甘蓝型油菜中存在3个具有功能的ALS酶(ALS1、ALS2、ALS3),对应的基因为BnALS1、BnALS2、BnALS3(相应的NCBI登录号为Z11524、Z11525、Z11526)。为明确哪一ALS基因何种突变是K5突变体获得对除草剂苯磺隆产生抗性的原因,申请人克隆了K5突变体的BnALS1、BnALS2、BnALS3三个基因,并对其序列完整性和与已公布序列的一致性进行验证。对此,根据已公布3个Topas甘蓝型油菜的ALS基因,设计引物分别用于克隆BnALS1、BnALS2、BnALS3;其中:
BnALS1(Bnals1544T)引物1:TCTCCTCTAACCATGGCGG和BnALS1(Bnals1544T)引物2:TCACCAGCTTCATCTCTCAGTAC。
BnALS2引物1:AGCAATTTCTCGCAACACTC和BnALS2引物2:AAATAAAGAGTGAAGTTTGCGT。
BnALS3引物1:CTCTCTCATCTAACCATGGC和BnALS3引物2:GGTCGACGATTACTGAAACTA。
使用植物基因组DNA提取试剂盒(CWBIO,新型植物基因组DNA提取试剂盒,Cat:CW0531A)提取野生型油菜(中双9号)与K5突变体油菜的叶片基因组DNA,使用KOD-FX-NEO(东洋纺,Code Number:KFX-201,PCR体系按照说明书配置)分别克隆野生型油菜和K5突变体的BnALS1-BnALS3基因。PCR反应在BIORAD C1000Thermal cycler中进行,程序如下:94℃预变性3min;98℃变性10sec,58℃退火30sec,68℃延伸75sec,反应共35个循环;68℃复性7min。PCR产物经1%琼脂糖凝胶电泳分离后,使用胶回收试剂盒(AXYGEN,DNA Gel Extraction Kit,Case#:0298)回收相应的目的片段;纯化PCR产物经加A后,连入pMDTM19T克隆载体中(Takara,Cat#:6013),通过热激转化进入大肠杆菌DH5α感受态细胞。挑取单克隆进行菌落PCR,将阳性克隆送往上海生工生物有限公司进行测序鉴定。测序结果表明,甘蓝型油菜K5突变体BnALS1基因与野生型油菜的BnALS1在+544碱基处存在一个SNP的差异,碱基由C突变为T(SEQ ID NO.1),通过对已公布Topas的BnALS1序列、中双9号BnALS1以及K5突变体BnALS1序列进行比对,确证K5突变体BnALS1氨基酸序列(SEQ ID NO.2)在182位的脯氨酸由丝氨酸替换(图2)。而K5突变体的BnALS2和BnALS3基因与野生型的对应基因序列一致。
(二)抗性基因Bnals1544T的功能验证
1、表达载体构建
在(一)的BnALS1引物1和引物2的5’端分别加上XmaI和BstEII酶切位点,形成新的BnALS1引物1’和BnALS1引物2’,其中:
BnALS1引物1’:CCCCCGGGTCTCCTCTAACCATGGCGG
BnALS1引物2’:GGGTCACCTCACCAGCTTCATCTCTCAGTAC
以(一)中测序存在的阳性克隆菌株为模板,利用新的BnALS1引物1’和BnALS1引物2’进行PCR扩增,经克隆测序验证,摇菌并提取带有Bnals1544T基因的阳性质粒p19T-Bnals1544T(AXYGEN,Plasmid Miniprep Kit,Case#:42)。通过XmaI(Thermo Scientific,code:#ER0171)和BstEII(Thermo Scientific,code:#ER0391)双酶切与T4 DNA连接酶(Takara,T4 DNA Ligase,Code:2011A)将目的片段连入经过改造的pCAMBIA3301植物表达载体(在EcoRI和SacI之间连入一个CaMV35S启动子)中,连接产物经热激转化进入DH5α大肠杆菌感受态细胞,挑取单克隆菌落进行菌落PCR,得到的阳性菌株经过摇菌、提取质粒,获得带有Bnals1544T的重组表达载体(pC3301-35S::Bnals1544T)。使用热激法将pC3301-35S::Bnals1544T转入农杆菌GV3101感受态细胞,又经菌落PCR,得到带有目的基因Bnals1544T的重组农杆菌菌株GV-pC3301-35S::Bnals1544T。
2、Bnals1544T转拟南芥功能验证
通过农杆菌介导的浸花法(Zhang et al.2006,Nature Protocols 1:641-646)侵染野生型哥伦比亚拟南芥,将Bnals1544T导入拟南芥中。T0代拟南芥种子种于培养钵中,待所有种子均发芽后,叶面喷施有效浓度为万分之一的草铵膦,每48h喷施一次,共喷施3次,筛选出12株阳性转基因植株。提取阳性转基因植株的叶片基因组DNA,使用BnALS1引物1和引物2进行PCR检测,所用20μl PCR反应体系为:10μl 2×Es taq MasterMix(CWBIO,Cat:CW0690),7μl双蒸水,引物1’与引物2’各0.5μl;2μl的DNA模板;反应程序为:94℃预变性3min;94℃变性30sec,58℃退火30sec,72℃延伸1min,共30个循环;72℃复性5min,PCR鉴定结果如图3所示,鉴定结果表明,其中有3株(NO.8,NO.11,NO.12)为假阳性植株,其余12株为转基因成功的阳性植株。
在6片真叶时,对阳性转基因植株喷施2μg/ml的苯磺隆溶液(野生型拟南芥致死浓度为0.1μg/ml),得到3株对苯磺隆具有明显抗性的转基因植株(图4)。结果表明,抗性基因Bnals1544T是植株获得对苯磺隆抗性的原因。
核苷酸或氨基酸序列表
<110>西北农林科技大学大学
<120>一种甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因及其应用
<160>
<210>1
<211>1968
<212>SEQ ID NO.1
<213>
<220>
<400>1
ATGGCGGCGGCAACATCGTCTTCTCCGATCTCCTTAACCGCTAAACCTTCTTCCAAATCCCCTCTACCCATTTCCAGATTCTCCCTTCCCTTCTCCTTAACCCCACAGAAAGACTCCTCCCGTCTCCACCGTCCTCTCGCCATCTCCGCCGTTCTCAACTCACCCGTCAATGTCGCACCTCCTTCCCCTGAAAAAACCGACAAGAACAAGACTTTCGTCTCCCGCTACGCTCCCGACGAGCCCCGCAAGGGTGCTGATATCCTCGTCGAAGCCCTCGAGCGTCAAGGCGTCGAAACCGTCTTTGCTTATCCCGGAGGTGCTTCCATGGAGATCCACCAAGCCTTGACTCGCTCCTCCACCATCCGTAACGTCCTTCCCCGTCACGAACAAGGAGGAGTCTTCGCCGCCGAGGGTTACGCTCGTTCCTCCGGCAAACCGGGAATCTGCATAGCCACTTCGGGTCCCGGAGCTACCAACCTCGTCAGCGGGTTAGCAGACGCGATGCTTGACAGTGTTCCTCTTGTCGCCATTACAGGACAGGTCTCTCGCCGGATGATCGGTACTGACGCCTTCCAAGAGACACCAATCGTTGAGGTAACGAGGTCTATTACGAAACATAACTATTTGGTGATGGATGTTGATGACATACCTAGGATCGTTCAAGAAGCTTTCTTTCTAGCTACTTCCGGTAGACCCGGACCGGTTTTGGTTGATGTTCCTAAGGATATTCAGCAGCAGCTTGCGATTCCTAACTGGGATCAACCTATGCGCTTACCTGGCTACATGTCTAGGTTGCCTCAGCCTCCGGAAGTTTCTCAGTTAGGTCAGATCGTTAGGTTGATCTCGGAGTCTAAGAGGCCTGTTTTGTACGTTGGTGGTGGAAGCTTGAACTCGAGTGAAGAACTGGGGAGATTTGTCGAGCTTACTGGGATCCCCGTTGCGAGTACTTTGATGGGGCTTGGCTCTTATCCTTGT AACGATGAGTTGTCCCTGCAGATGCTTGGCATGCACGGGACTGTGTATGCTAACTACGCTGTGGAGCATAGTGATTTGTTGCTGGCGTTTGGTGTTAGGTTTGATGACCGTGTCACGGGAAAGCTCGAGGCTTTCGCTAGCAGGGCTAAAATTGTGCACATAGACATTGATTCTGCTGAGATTGGGAAGAATAAGACACCTCACGTGTCTGTGTGTGGTGATGTAAAGCTGGCTTTGCAAGGGATGAACAAGGTTCTTGAGAACCGGGCGGAGGAGCTCAAGCTTGATTTCGGTGTTTGGAGGAGTGAGTTGAGCGAGCAGAAACAGAAGTTCCCTTTGAGCTTCAAAACGTTTGGAGAAGCCATTCCTCCGCAGTACGCGATTCAGATCCTCGACGAGCTAACCGAAGGGAAGGCAATTATCAGTACTGGTGTTGGACAGCATCAGATGTGGGCGGCGCAGTTTTACAAGTACAGGAAGCCGAGACAGTGGCTGTCGTCATCAGGCCTCGGAGCTATGGGTTTTGGACTTCCTGCTGCGATTGGAGCGTCTGTGGCGAACCCTGATGCGATTGTTGTGGATATTGACGGTGATGGAAGCTTCATAATGAACGTTCAAGAGCTGGCCACAATCCGTGTAGAGAATCTTCCTGTGAAGATACTCTTGTTAAACAACCAGCATCTTGGGATGGTCATGCAATGGGAAGATCGGTTCTACAAAGCTAACAGAGCTCACACTTATCTCGGGGACCCGGCAAGGGAGAACGAGATCTTCCCTAACATGCTGCAGTTTGCAGGAGCTTGCGGGATTCCAGCTGCGAGAGTGACGAAGAAAGAAGAACTCCGAGAAGCTATTCAGACAATGCTGGATACACCAGGACCATACCTGTTGGATGTGATATGTCCGCACCAAGAACATGTGTTACCGATGATCCCAAGTGGTGGCACTTTCAAAGATGTAATAACAGAAGGGGATGGTCGCACTAAGTACTGA
<210>2
<211>655
<212>SEQ ID NO.2
<213>
<220>
<400>2
MAAATSSSPISLTAKPSSKSPLPISRFSLPFSLTPQKDSSRLHRPLAISAVLNSPVNVAPPSPEKTDKNKTFVSRYAPDEPRKGADILVEALERQGVETVFAYPGGASMEIHQALTRSSTIRNVLPRHEQGGVFAAEGYARSSGKPGICIATSGPGATNLVSGLADAMLDSVPLVAITGQVSRRMIGTDAFQETP IVEVTRSITKHNYLVMDVDDIPRIVQEAFFLATSGRPGPVLVDVPKDIQQQLAIPNWDQPMRLPGYMSRLPQPPEVSQLGQIVRLISESKRPVLYVGGGSLNSSEELGRFVELTGIPVASTLMGLGSYPCNDELSLQMLGMHGTVYANYAVEHSDLLLAFGVRFDDRVTGKLEAFASRAKIVHIDIDSAEIGKNKTPHVSVCGDVKLALQGMNKVLENRAEELKLDFGVWRSELSEQKQKFPLSFKTFGEAIPPQYAIQILDELTEGKAIISTGVGQHQMWAAQFYKYRKPRQWLSSSGLGAMGFGLPAAIGASVANPDAIVVDIDGDGSFIMNVQELATIRVENLPVKILLLNNQHLGMVMQWEDRFYKANRAHTYLGDPARENEIFPNMLQFAGACGIPAARVTKKEELREAIQTMLDTPGPYLLDVICPHQEHVLPMIPSGGTFKDVITEGDGRTKY
<210>3
<211>19
<212>Bnals1544T的引物1
<213>
<220>
<400>3
TCTCCTCTAACCATGGCGG
<210>4
<211>23
<212>Bnals1544T的引物2
<213>
<220>
<400>4
TCACCAGCTTCATCTCTCAGTAC
<210>5
<211>20
<212>BnALS2引物1
<213>
<220>
<400>5
AGCAATTTCTCGCAACACTC
<210>6
<211>22
<212>BnALS2引物2
<213>
<220>
<400>6
AAATAAAGAGTGAAGTTTGCGT
<210>7
<211>20
<212>BnALS3引物1
<213>
<220>
<400>7
CTCTCTCATCTAACCATGGC
<210>8
<211>21
<212>BnALS3引物2
<213>
<220>
<400>8
GGTCGACGATTACTGAAACTA
<210>9
<211>27
<212>BnALS1引物1’
<213>
<220>
<400>9
CCCCCGGGTCTCCTCTAACCATGGCGG
<210>10
<211>31
<212>BnALS2引物2’
<213>
<220>
<400>10
GGGTCACCTCACCAGCTTCATCTCTCAGTAC。
核苷酸或氨基酸序列表
<110> 西北农林科技大学
<120> 一种甘蓝型油菜抗磺酰脲类除草剂苯磺隆的基因及应用
<160>
<210> 1
<211> 1968
<212> SEQ ID NO.1
<213>
<220>
<400> 1
ATGGCGGCGGCAACATCGTCTTCTCCGATCTCCTTAACCGCTAAACCTTCTTCCAAATCCCCTCTACCCATTT
CCAGATTCTCCCTTCCCTTCTCCTTAACCCCACAGAAAGACTCCTCCCGTCTCCACCGTCCTCTCGCCATCTC
CGCCGTTCTCAACTCACCCGTCAATGTCGCACCTCCTTCCCCTGAAAAAACCGACAAGAACAAGACTTTCGTC
TCCCGCTACGCTCCCGACGAGCCCCGCAAGGGTGCTGATATCCTCGTCGAAGCCCTCGAGCGTCAAGGCGTCG
AAACCGTCTTTGCTTATCCCGGAGGTGCTTCCATGGAGATCCACCAAGCCTTGACTCGCTCCTCCACCATCCG
TAACGTCCTTCCCCGTCACGAACAAGGAGGAGTCTTCGCCGCCGAGGGTTACGCTCGTTCCTCCGGCAAACCG
GGAATCTGCATAGCCACTTCGGGTCCCGGAGCTACCAACCTCGTCAGCGGGTTAGCAGACGCGATGCTTGACA
GTGTTCCTCTTGTCGCCATTACAGGACAGGTCTCTCGCCGGATGATCGGTACTGACGCCTTCCAAGAGACACC
AATCGTTGAGGTAACGAGGTCTATTACGAAACATAACTATTTGGTGATGGATGTTGATGACATACCTAGGATC
GTTCAAGAAGCTTTCTTTCTAGCTACTTCCGGTAGACCCGGACCGGTTTTGGTTGATGTTCCTAAGGATATTC
AGCAGCAGCTTGCGATTCCTAACTGGGATCAACCTATGCGCTTACCTGGCTACATGTCTAGGTTGCCTCAGCC
TCCGGAAGTTTCTCAGTTAGGTCAGATCGTTAGGTTGATCTCGGAGTCTAAGAGGCCTGTTTTGTACGTTGGT
GGTGGAAGCTTGAACTCGAGTGAAGAACTGGGGAGATTTGTCGAGCTTACTGGGATCCCCGTTGCGAGTACTT
TGATGGGGCTTGGCTCTTATCCTTGTAACGATGAGTTGTCCCTGCAGATGCTTGGCATGCACGGGACTGTGTA
TGCTAACTACGCTGTGGAGCATAGTGATTTGTTGCTGGCGTTTGGTGTTAGGTTTGATGACCGTGTCACGGGA
AAGCTCGAGGCTTTCGCTAGCAGGGCTAAAATTGTGCACATAGACATTGATTCTGCTGAGATTGGGAAGAATA
AGACACCTCACGTGTCTGTGTGTGGTGATGTAAAGCTGGCTTTGCAAGGGATGAACAAGGTTCTTGAGAACCG
GGCGGAGGAGCTCAAGCTTGATTTCGGTGTTTGGAGGAGTGAGTTGAGCGAGCAGAAACAGAAGTTCCCTTTG
AGCTTCAAAACGTTTGGAGAAGCCATTCCTCCGCAGTACGCGATTCAGATCCTCGACGAGCTAACCGAAGGGA
AGGCAATTATCAGTACTGGTGTTGGACAGCATCAGATGTGGGCGGCGCAGTTTTACAAGTACAGGAAGCCGAG
ACAGTGGCTGTCGTCATCAGGCCTCGGAGCTATGGGTTTTGGACTTCCTGCTGCGATTGGAGCGTCTGTGGCG
AACCCTGATGCGATTGTTGTGGATATTGACGGTGATGGAAGCTTCATAATGAACGTTCAAGAGCTGGCCACAATCC
GTGTAGAGAATCTTCCTGTGAAGATACTCTTGTTAAACAACCAGCATCTTGGGATGGTCATGCAATGGGAAGATCG
GTTCTACAAAGCTAACAGAGCTCACACTTATCTCGGGGACCCGGCAAGGGAGAACGAGATCTTCCCTAACATGCT
GCAGTTTGCAGGAGCTTGCGGGATTCCAGCTGCGAGAGTGACGAAGAAAGAAGAACTCCGAGAAGCTATTCAGACA
ATGCTGGATACACCAGGACCATACCTGTTGGATGTGATATGTCCGCACCAAGAACATGTGTTACCGATGATCC
CAAGTGGTGGCACTTTCAAAGATGTAATAACAGAAGGGGATGGTCGCACTAAGTACTGA(1968 bp)
<210> 2
<211> 655
<212> SEQ ID NO.2
<213>
<220>
<400> 2
MAAATSSSPISLTAKPSSKSPLPISRFSLPFSLTPQKDSSRLHRPLAISAVLNSPVNVAPPSPEKTDKNKTFV
SRYAPDEPRKGADILVEALERQGVETVFAYPGGASMEIHQALTRSSTIRNVLPRHEQGGVFAAEGYARSSGKP
GICIATSGPGATNLVSGLADAMLDSVPLVAITGQVSRRMIGTDAFQETPIVEVTRSITKHNYLVMDVDDIPRI
VQEAFFLATSGRPGPVLVDVPKDIQQQLAIPNWDQPMRLPGYMSRLPQPPEVSQLGQIVRLISESKRPVLYVGGGS
LNSSEELGRFVELTGIPVASTLMGLGSYPCNDELSLQMLGMHGTVYANYAVEHSDLLLAFGVRFDDRVTGKLEAFA
SRAKIVHIDIDSAEIGKNKTPHVSVCGDVKLALQGMNKVLENRAEELKLDFGVWRSELSEQKQKFPLSFKTFGEAI
PPQYAIQILDELTEGKAIISTGVGQHQMWAAQFYKYRKPRQWLSSSGLGAMGFGLPAAIGASVANPDAIVVDIDGD
GSFIMNVQELATIRVENLPVKILLLNNQHLGMVMQWEDRFYKANRAHTYLGDPARENEIFPNMLQFAGACGIPAAR
VTKKEELREAIQTMLDTPGPYLLDVICPHQEHVLPMIPSGGTFKDVITEGDGRTKY(655 aa)
<210> 3
<211> 19
<212> Bnals1544T的引物1
<213>
<220>
<400>3
TCTCCTCTAACCATGGCGG
<210> 4
<211> 23
<212> Bnals1544T的引物2
<213>
<220>
<400> 4
TCACCAGCTTCATCTCTCAGTAC
<210> 5
<211> 20
<212> BnALS2引物1
<213>
<220>
<400> 5
AGCAATTTCTCGCAACACTC
<210> 6
<211> 22
<212> BnALS2引物2
<213>
<220>
<400> 6
AAATAAAGAGTGAAGTTTGCGT
<210> 7
<211> 20
<212> BnALS3引物1
<213>
<220>
<400> 7
CTCTCTCATCTAACCATGGC
<210> 8
<211> 21
<212> BnALS3引物2
<213>
<220>
<400> 8
GGTCGACGATTACTGAAACTA
<210> 9
<211> 27
<212> BnALS1引物1’
<213>
<220>
<400> 9
CCCCCGGGTCTCCTCTAACCATGGCGG
<210> 10
<211> 31
<212> BnALS2引物2’
<213>
<220>
<400> 10
GGGTCACCTCACCAGCTTCATCTCTCAGTAC
- 还没有人留言评论。精彩留言会获得点赞!