包含163G>C突变的FKBP5基因片段、所编码的蛋白质片段及其应用的制作方法
本发明属于生物医学领域,涉及fkbp5基因的突变。
背景技术:
:paget骨病是一种局限性骨组织异常重构的慢性代谢性骨病,主要累及年龄长于55岁的老年人(1)。paget骨病的流行特点具有显著的地域差异性,在英国、澳大利亚及北美等一些地区具有较高发病率,是仅次于骨质疏松的第二大骨病,然而在非洲、亚洲地区却极为少见(2)。paget骨病病因仍不清楚,普遍认为遗传因素在其发生发展中具有重要作用。paget骨病易感基因主要包括:sqstm1、tnfrsf11a、tnfrsf11b、optn、vcp、csf1以及dcstamp等(3-6)。其中,sqstm1基因突变被认为是paget骨病的致病基因突变,迄今为止已发现28个不同的sqstm1突变存在于约1/3的pdb家系和9%的散发型pdb患者中(7)。破骨细胞功能异常是paget骨病最主要的病理改变。患者病变部位破骨细胞数目增多、体积增大、细胞核数量增多(8)。患者外周血来源的单核细胞对破骨细胞诱导因子rankl及1,25-(oh)2维生素d3敏感性增加,说明paget骨病患者体内破骨细胞分化及功能异常(9)。破骨细胞由骨髓内单核/巨噬细胞经由rankl因子诱导分化而来,其分化过程中主要活化的信号通路包括nf-κb、akt信号通路(10)。因此,异常的nf-κb、akt信号通路调控可能参与paget骨病发生。fkbp5基因位于第6号染色体上,包括12个外显子,基因全长154999bp。fkbp5蛋白(fk506结合蛋白51,fkbp5)是一种具有多种生物学功能的免疫亲和素,属于fkbp家族中的大分子免疫亲和素,该家族因为能够与免疫抑制剂fk506结合而得名。fkbp5蛋白共有三个结构域,包括n-末端的fk1、fk2(fkbp12样结构域,fk)结构域以及c-末端的tpr(tetratricopeptiderepeat)结构域。其中,fk1结构能够与fk506等免疫抑制剂结合,并且具有肽辅酰胺异构酶活性(ppiase)。fk2结构域的结构与fk1类似,但是不具备免疫抑制药物结合能力以及ppiase活性。位于c-末端的trp结构能够与hsp90蛋白结合,与之构成类固醇激素受体复合物的重要伴侣分子,调控类固醇激素受体活性。fkbp5蛋白生物学功能复杂多样,其在类固醇激素受体调节、压力相关精神疾病发生、癌症发生与放化疗敏感性、自噬等生理及病理过程中均发挥重要作用。信号通路调控方面,fkbp5作为ikkα重要的伴侣分子,参与iκb磷酸化及降解过程,从而增加nf-κb信号通路活性(11);在akt信号通路中,fkbp5作为桥联蛋白参与akt与phlpp(ph域和富亮氨酸重复蛋白磷酸酶)之间的相互作用,促进phlpp对aktser473位点的去磷酸化作用,从而降低akt磷酸化水平(12)。技术实现要素:本申请发明人研究发现,fkbp5基因的突变与paget骨病的发病相关。基于此研究,本申请的目的是提供一种新的与paget骨病相关的基因突变,以用于检测paget骨病。在第一个方面,本发明提供核酸片段,所述核酸片段包含对应于seqidno.1的第163位的核苷酸,且包含seqidno.1所示的核苷酸序列中的至少10个连续核苷酸,其中第163位的核苷酸是c;或者所述核酸片段包含对应于seqidno.2的第479位的核苷酸,且包含seqidno.2所示的核苷酸序列中的至少10个连续核苷酸,其中第479位的核苷酸是c;或者所述核酸片段包含对应于seqidno.3的第1001位的核苷酸,且包含seqidno.3所示的核苷酸序列中的至少10个连续核苷酸,其中第1001位的核苷酸是c;或者所述核酸片段为上述核酸片段的反向互补序列。优选地,所述核酸片段的长度可以是10-100、101-200、201-500或501-1000个核苷酸。优选地,所述核酸片段的长度是10-20、21-30、31-40、41-50、51-60、61-100或101-300个核苷酸。所述突变位点可以位于所述核酸片段的任何位置。优选地,所述核酸片段是seqidno.1所示的序列。优选地,所述核酸片段是seqidno.2所示的序列。优选地,所述核酸片段是seqidno.3所示的序列。优选地,所述核酸片段可以是seqidno.7或8所示的序列。本发明的第二个方面是提供用于检测和/或分析本发明的单碱基突变的引物或试剂盒,所述引物能够扩增单碱基突变,所述单碱基对应于seqidno.1的第163位或seqidno.2的第479位或seqidno.3的第1001位;所述试剂盒包含所述引物。优选地,所述引物序列如seqidno.5和seqidno.6所示;所述试剂盒包含序列为seqidno.5和seqidno.6的引物。本发明的第三个方面是提供本发明的核酸片段在制备检测fkbp5基因突变的制剂中的应用;或者本发明的核酸片段用作检测fkbp5基因突变的检验标志物的应用。本发明的第四个方面是提供fkbp5蛋白或其片段或变异体,所述蛋白序列为seqidno.4所示的序列;所述片段或变异体包含对应于seqidno.4的第55位的亮氨酸,并且包含seqidno.4所示的氨基酸序列的至少10个连续氨基酸,如10-20、21-50或51-100个氨基酸。本发明的第五个方面是提供分析核酸的方法,所述方法包括分析待测样品中的包含对应于seqidno.1的序列的核酸中对应于第163位的核苷酸或分析待测样品中的包含对应于seqidno.2的序列的核酸中对应于第479位的核苷酸或分析待测样品中的包含对应于seqidno.3的序列的核酸中对应于第1001位的核苷酸。在一种实施方式中,所述方法可以是限制性片段长度多态性分析(rflp)。本领域技术人员可以根据本
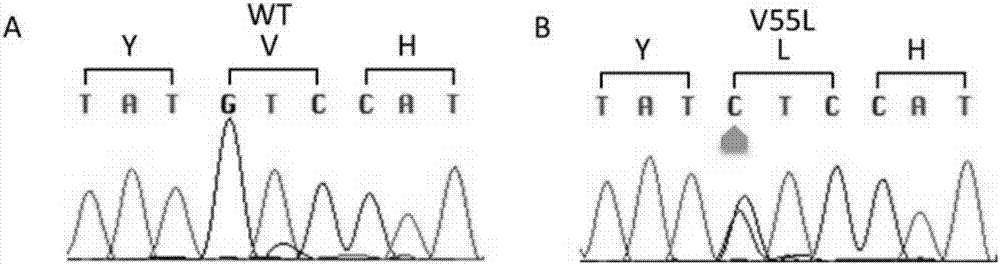
发明内容设计实验以分析对应于seqidno.1的序列的核酸中的第163位的核苷酸或对应于seqidno.2的序列的核酸中的第479位的核苷酸是否为c或对应于seqidno.3的序列的核酸中的第1001位的核苷酸是否为c。在另一种实施方式中,所述方法可以是测序法,包括分离并测定来自基因组dna或rna的核酸序列,分析其中包含对应于seqidno.1的序列的核酸中的对应于第163位的核苷酸或包含对应于seqidno.2的序列的核酸中的对应于第479位的核苷酸或包含对应于seqidno.3的序列的核酸中的对应于第1001位的核苷酸是否为c。测序法可以是本领域已知的任何可用的测序方法。测序引物可以根据本领域技术人员的常识进行设计,如在待检测位点的上下游适当位置处设计引物,以扩展包含该待测位点的片段,从而判断该位点的核苷酸。也可以采用本发明的寡核苷酸作为引物序列。在另一种实施方式中,所述方法是利用探针杂交的方法,特异性地鉴定检测样品中包含对应于seqidno.1的序列的核酸中的对应于第163位的核苷酸或包含对应于seqidno.2的序列的核酸中对应于第479位的核苷酸或包含对应于seqidno.3的序列的核酸中对应于第1001位的核苷酸是否为c;所述方法中采用的探针是本发明的寡核苷酸。例如,从待测样品中分离出核酸,在允许探针与核酸中可能存在的特异性靶序列杂交的条件下使探针与核酸接触;可被检测的杂交可以通过使用由可检测的试剂标记过的探针来实现;例如,用放射性同位素、荧光染料或能催化形成可检测产物的酶来标记探针。标记探针、用标记探针检测样品中是否存在靶序列的方法都是本领域技术人员所熟知的。本发明中,所述样品可以是任何包含核酸的样品,如血液;优选所述样品来自于人。所述核酸可以是dna或rna,优选为基因组dna。本发明的分析核酸的方法可以以dna或rna为目标物。本领域技术人员可知,当以dna为检测目标物时,分析待测样品中的包含对应于seqidno.3的序列的核酸中对应于第1001位的核苷酸,所使用的探针或引物根据seqidno.3的序列设计;当以rna为检测目标物时,分析待测样品中的包含对应于seqidno.2的序列的核酸中对应于第479位的核苷酸,所使用的探针或引物根据seqidno.2的序列设计。本发明提供了包含新的单碱基突变的fkbp5基因和其编码序列,该基因在对应于seqidno.1的第163位核苷酸由g突变为c(163g>c),从而引起其编码的氨基酸由缬氨酸突变为亮氨酸,即对应于seqidno.4的第55位的亮氨酸。该突变的fkbp5蛋白增高了aktser473磷酸化水平,增加了破骨细胞能力,导致paget骨病的形成。由于paget骨病为迟发病,通常累及年龄长于55岁者,因而本发明为paget骨病的提前诊断提供了新的标志物,具有很强的临床指导意义。附图说明图1:家族聚集性paget骨病病例家系图;其中圆代表女性,方框代表男性;图2:fkbp5基因突变位点测序图;a:显示野生型fkbp5基因cdna第160-168位碱基序列。b:显示突变型fkbp5基因cdna第160-168位碱基序列,图中箭头所示为突变位点碱基,测序峰图显示该突变为杂合子突变;图3:fkbp5真核表达载体图谱;图4:转染不同质粒的hek293细胞的相对荧光素酶活性;图5:nf-κb核转位水平检测结果;图6:aktser473位点磷酸化水平检测;其中,a所示为hek293细胞转染野生型与突变型fkbp5后,与转染空载体的对照组相比fkbp5蛋白质表达水平明显增高,证明质粒转染成功,fkbp5过表达;b为各组akt磷酸化检测wb显影条带图;c为wb条带灰度值paktser473与总akt比值,显示aktser473位点磷酸化程度。野生组低于突变组与对照组,突变组与对照组之间无显著差异;图7:fkbp5v55l敲入小鼠打靶及构建策略简图;图8:小鼠破骨细胞trap染色图;图9:小鼠破骨细胞数目统计图;图10:陷窝吸收实验结果图;其中,a:显示野生组与突变组小鼠破骨细胞在牛骨切片上形成的吸收陷窝,图中蓝染部分即为破骨细胞骨吸收陷窝;b:两组细胞形成的陷窝在低倍镜(40×)视野下拍照,图片中吸收陷窝面积使用imageproplus软件进行计算;所得每组每视野下平均吸收面积作图,并进行统计学分析;图11:小鼠股骨micro-ct检测骨参数分析结果;图12:小鼠股骨micro-ct检测2d平面图;图13:小鼠股骨远端micro-ct检测3d重构图;图14:小鼠测序鉴定序列图,其中,与其cdna序列第163位点相对应的位置:野生鼠测序峰图显示碱基为g,杂合鼠为g/c双峰重叠,而纯合突变鼠峰图为碱基c;图15:琼脂糖凝胶鉴定小鼠基因型结果图。具体实施方式下面结合具体实施方式说明本发明,但本发明的内容不限于此。在以下实施例中,如无特别说明,所使用的试剂的仪器都是本领域常规试剂和仪器,可以从商购途径获得;所使用的方法为本领域常规方法,本领域技术人员根据本发明的描述和实验目的,可以毫无疑问地知道如何进行所述实验并实现实验目的。如无其它说明,本发明的“核酸片段”由核苷酸或其类似物组成,可以是dna、rna或其类似物的片段;可以是单链或双链;可以是天然的(如基因组的)或合成的。本发明中,“突变”指在检测的基因,即fkbp5基因中存在与野生型fkbp5基因序列不同的核苷酸位点。“突变位点”指碱基发生突变的位置。在本发明中,所述突变位点是对应于seqidno.1所示序列的第163位或seqidno.2所示序列中的第479位或seqidno.3所示序列中的第1001位。本
发明内容涉及fkbp5基因的非同义突变。由于该突变位点位于基因的编码序列中,因此,本领域技术人员可知,所述突变位点既可以在基因组dna中表现,也可以在编码序列(即cds)中表现。本领域技术人员根据待检测样品,可以在基因组dna或mrna水平上对该突变位点进行检测。本申请中,seqidno.1是具有上述突变的fkbp5的cds序列,其中第163位涉及该突变(g>c);seqidno.2是具有所述突变位点的fkbp5的mrna序列,其中第479位是本发明涉及的突变位点;seqidno.3是具有所述突变位点的fkbp5基因所在的基因组的部分序列,其中第1001位是本发明涉及的突变位点。本领域技术人员可知,在本文中,对应于seqidno.1的第163位位点、对应于seqidno.2的第479位位点和对应于seqidno.3的第1001位位点同义互用。本发明中,核苷酸和氨基酸的缩写采用本领域公知的缩写方式。本发明的内容是基于fkbp5基因的新的单碱基突变位点。所述突变位点是位于fkbp5基因的编码区内,对应于seqidno.1的第163位,该位点由野生型的g突变为c;由该突变的fkbp5基因编码的蛋白的第55位由缬氨酸突变为亮氨酸(v55l)。实施例1:人fkbp5基因突变位点的获得从济南市第三人民医院得一例中国汉族人群家族聚集性paget骨病病例,家系图谱见图1,家族成员均知情同意本研究。家系中共有3位患者,分别为ⅱ3、ⅱ6、ⅱ11。根据患者骨骼系统影像学检查结果、血清学检查结果、临床症状及体征可以明确诊断为paget骨病。抽取患者及其他家系成员的外周血全血,使用qiagen全血小量dna提取试剂盒提取基因组dna,进行基因突变检测分析。1、paget骨病易感基因测序分析针对已知的paget易感基因,包括sqstm1、tnfrsf11a、tnfrsf11b、optn、vcp、csf1以及dcstamp,设计引物,使用sanger测序分析所有家系成员中这些基因的外显子区域,并与已鉴定的与paget骨病相关的snp进行比较。这些基因的测序引物见后面。sanger测序分析结果显示,全部检测到的snp位点中均无位于外显子的非同义突变,排除这些snp致病的可能,该家系不存在已知的paget骨病易感基因突变,这提示可能存在新的基因突变参与paget骨病发生。检测到的snp位点见表1。表1:家系中检测到的paget骨病易感基因snp位点注:snp定位于外显子;snp定位于5’-utr;§:snp定位于剪切位点。2、外显子组测序分析选取全部三位患者(ⅱ3、ⅱ6、ⅱ11)及患者的正常兄妹(ⅱ4、ⅱ8)的dna为样本,进行外显子组测序分析。外显子组测序由北京博莱明创公司完成。该公司所使用的外显子富集试剂盒为美国agilenttechnologies公司的agilentsure-selecthumanallexonkit,测序平台为illuminahiseq2000测序平台。外显子组测序所检测到的所有单核苷酸变异(snv),按照下列条件筛选候选paget骨病致病基因突变:a)不存在于已知的snp数据库中,即non-dbsnp;b)存在于所有三位患者基因组dna当中;c)不存在于正常对照的基因组dna当中;d)存在于基因外显子序列中的非同义单核苷酸突变。根据上述筛选条件所得单核苷酸突变,进一步在exomeaggregationconsortium(exac)数据库中检索,并使用polyphen-2预测突变功能。排除已知的以及功能预测结果为良性的突变。snp注释结果显示:5例测序样本共得到245982个单核苷酸变异(snv),其中,不存在于已知的snp数据库中的snv(non-dbsnpsnv)共有82676个,同时存在于三位患者当中且不存在于两位对照当中的non-dbsnpsnv,共有193个。最终,排除所有同义突变以及基因编码区之外的non-dbsnpsnv,共得到5个位于不同基因的non-dbsnpsnv与疾病共分离,包括fkbp5(c.163g>c)、acbd4(c.4g>t)、myo7b(c.772a>t)、akna(c.4196a>g)、ccl4l1(c.197g>c),见表2。表2:家系中与疾病共分离的五个基因突变在exac数据库中检索上述5个基因突变,其中三个突变存在于该数据库中,包括:acbd4(c.4g>t)、myo7b(c.772a>t)及ccl4l1(c.197g>c)。fkbp5(c.163g>c)及akna(c.4196a>g)未见报道。ployphen-2预测结果显示:fkbp5(c.163g>c)基因突变极有可能影响蛋白功能(probablydamaging:0.992);而akna(c.4196a>g)基因突变则极有可能为良性突变,不影响蛋白功能(benign:0.080)。fkbp5基因位于6p21.31,位于一个pdb易感位点(pdb1)内,发现的163g>c突变位于fkbp5基因第4外显子内。其编码的fkbp5蛋白属于免疫亲和素家族的一员,生物学功能与破骨细胞分化发育紧密相关。fkbp5参与多种生物学过程,包括调节类固醇激素受体活化,促进微管系统稳定,促进nf-κb信号通路活化,抑制akt磷酸化等。其中,nf-κb以及akt信号通路均参与调节破骨细胞分化发育。破骨细胞异常活化是pdb重要的病理改变,因此fkbp5突变可能为pdb发病的致病基因突变。实施例2:fkbp5基因突变验证为在家系中验证该基因突变以及确认该突变是否为汉族人群中未经报道过的snp位点,对所有家系成员以及200例正常对照的fkbp5基因第4外显子包含突变位点的基因序列进行测序分析。1、含突变位点163g>c的片段pcr以所有家系成员以及200名正常对照的基因组dna为模板,扩增包含有fkbp5c.163g>c基因突变位点的基因片段。扩增引物片段为:上游5’-attaaaccaatgaacaggat-3’(seqidno.5);下游5’-cacaactcaaaaaaataccc-3’(seqidno.6);产物长度:446bp。pcr扩增体系如下:takaraextaqhs(5u/μl)0.1μl10×extaq缓冲液2μldntp混合物(每种2.5mm)2μldna100ng引物(上下游引物混合物,各10μm,)2μlddh2o补充到20μl充分混匀后,按照以下pcr循环条件,扩增目的片段:反应结束后,将pcr产物取出,4℃冰箱保存,准备纯化。2、pcr产物纯化1)向96孔pcr反应板内的20μlpcr产物中依次加入5μl5mnacl溶液以及25μl22%peg8000,封膜,充分混匀后瞬时离心,4℃静置大于30min;2)设置低温高速离心机,在4℃条件下,以2900×g离心力离心96孔pcr反应板内的混合物30分钟;3)弃掉板内上清液,将96孔板倒置,以小于180×g的离心力瞬时离心96孔板,去除板内残余液体;4)向板孔内加入80μl70%乙醇溶液,封膜;5)在4℃条件下,以2900×g离心力离心96孔板15min;6)弃掉上清,以小于180×g离心力扣板离心,弃掉板内残留液体;7)重复步骤4-6,再次洗涤沉淀;8)96孔板室温干燥15min,放置在4℃冰箱保存备用。3、纯化产物测序pcr将纯化的pcr产物取出,每孔加入5μl双蒸水溶解dna,作为测序pcr模板。测序pcr所用试剂盒是terminatorv3.1cycle测序试剂盒,反应体系如下:5×测序缓冲液1.16μlbigdye0.25μl测序引物(seqidno.5)(50μm)0.0625μldna模板1.5μlddh2o2.8275μl测序pcr反应条件:4、测序pcr产物纯化1)向测序pcr产物中依次加入7.5μl125μmedta溶液、7.5μl3mnaac溶液以及50μl无水乙醇,充分混匀后室温静置15分钟;2)设置低温高速离心机,4℃条件下,以2900×g离心力离心30分钟;3)弃掉板内上清,以小于180×g离心力扣板离心,去除板内残留液体;4)每孔加入80μl70%乙醇,封膜;5)4℃条件下,以2900×g离心力离心15分钟;6)弃掉上清,以小于180×g离心力扣板离心,去掉残留液体;7)重复步骤4-6,再洗涤dna沉淀一次;8)96孔板揭膜,在室温放置15分钟,干燥板内dna;9)每孔加入7μl甲酰胺,震板混匀,封膜;瞬时离心后,将板放入9700pcr仪,95℃变性5分钟,4℃放置大于2分钟;将板取出,准备测序。5、测序分析将准备好的测序dna产物放入abiprism310型全自动dna测序仪,进行测序。测序结果通过sequencher5.0软件比对。通过对家系成员以及200例正常对照进行fkbp5c.163g>c基因突变验证,证实三位患者均为c.163g>c杂合子突变,并且发现家系中有5位患者的后代(ⅲ3、ⅲ13、ⅲ15、ⅲ17、ⅳ2)携带有该基因突变,其基因型全部为gc杂合子(图2)。200例正常对照人群中并未检出该基因突变。实施例3:fkbp5基因突变对nf-κb、akt信号通路影响1、构建表达野生型及突变型fkbp5蛋白的真核表达载体pcdna3.1载体购自invitrogen(carlsbad,ca,usa)公司。通过设计引物,在fkbp5基因cds序列的5’端引入bamhi酶切位点,3’端引入ecori酶切位点。引物序列如下:上游5’-ggatccatgactactgatgaaggtg-3’;下游5’-gaattctcatacgtggccctcaggtt-3’。使用该引物,以野生型或突变型fkbp5基因cds序列为模板扩增目的片段。使用通用型dna纯化回收试剂盒(天根生化科技(北京)有限公司),回收并纯化pcr产物。纯化的pcr产物连接至pmd18-t(takara,tokyo,japan)连接载体上,经转化、涂板筛选、菌落pcr鉴定连接有野生型或突变型fkbp5基因cds序列的阳性克隆。扩增阳性pmd18-t载体并测序,序列正确的质粒使用bamhi和ecori(fermentas,thermofisherscientific,waltham,ma,usa)限制性内切酶酶切,并回收包含有野生型或突变型fkbp5基因cds序列的酶切产物。将所得酶切产物与事先使用bamhi和ecori限制性内切酶酶切为线性的pcdna3.1载体进行连接,经转化、涂板筛选、菌落pcr鉴定成功连接野生型或突变型fkbp5基因cds序列的pcdna3.1载体的阳性菌落。扩增阳性菌,提取质粒,即为实验所需能够表达野生型或突变型fkbp5蛋白的真核表达载体。真核表达载体的图谱如图3所示。2、荧光素酶报告基因检测将野生型pcdna3.1-fkbp5质粒、突变型pcdna3.1-fkbp5质粒、pcdna3.1质粒(空载体对照,购自invitrogen)分别与pgl4.32[luc2p/nf-κb-re/hygro]质粒(购自promega(madison,wi,usa))和prl-tk质粒(购自promega(madison,wi,usa))共转染到hek293细胞(购自美国atcc细胞库(manassas,va,usa))内,获得野生型组、突变型组和空载体对照组细胞,通过检测各转染组细胞的荧光素酶活性(luciferaseassay)反应nf-κb的转录启动活性。hek293细胞质粒转染及荧光素酶报告基因检测方法:将hek293细胞密度调整至2×105细胞/ml,接种到96孔全白酶标板中,每孔100μl细胞悬液,共接种9孔。野生型组(pcdna3.1-fkbp5(v55))、突变型组(pcdna3.1-fkbp5(l55))以及对照组(pcdna3.1)各设置3个复孔。细胞培养24小时后弃掉板内培养基,每孔加opti-mem转染培养基75μl。将质粒以及lipofectamine2000(invitrogen,carlsbad,ca,usa)转染试剂按照以下稀释比稀释(每孔用量):将稀释后的质粒和转染试剂混合,室温孵育10min后加入相应的孔内,放入孵箱培养4小时。转染4小时后换液,用dmem完全培养基继续培养细胞24小时。转染24小时后,向96孔板内加入tnf-α因子活化hek293细胞,终浓度20ng/ml。活化4小时候,裂解细胞并检测胞内荧光强度。使用dual-glo荧光素酶分析系统试剂盒检测荧光素酶活性,该试剂盒购自promega(madison,wi,usa)。简要操作步骤如下:将荧光素酶缓冲液全部加入到荧光素酶底物中,颠倒混匀,配制成荧光素酶试剂。计算stop&glo试剂用量,并将终止试剂和glo试剂以1:100稀释比稀释配制所需stop&glo试剂。每孔加入75μl荧光素酶试剂,室温孵育10min。使用enspiretm多模式板阅读仪读取并记录萤火虫荧光强度。继续向每孔中加入stop&glo试剂75μl,混匀后室温孵育10min。使用enspiretm多模式板阅读仪读取并记录海肾荧光强度。将所测萤火虫荧光值与海肾荧光值相比较,即为每一实验组的相对荧光素酶活性,反应各组nf-κb转录因子的转录启动活性。荧光素酶活性分析显示,野生型组、突变型组及空载体对照组hek293细胞的相对荧光素酶活性分别为:野生型组10.52±0.83,突变型组11.20±0.78,空载体对照组8.79±0.67。其中野生组与突变组之间相对荧光素酶活性无统计学差异,p=0.3624;野生组与突变组相对荧光素酶活性均高于对照组(野生组vs对照组:10.52±0.83vs8.79±0.67,p=0.0479;突变组vs对照组:11.20±0.78vs8.79±0.67,p=0.0152;图5)。说明转染fkbp5后,nf-κb转录启动活性增高。但是,v55l突变不影响fkbp5蛋白对nf-κb转录启动活性的调控作用。3、nf-κb核转位检测将野生型pcdna3.1-fkbp5质粒、突变型pcdna3.1-fkbp5质粒、pcdna3.1质粒分别转染到hek293细胞内,通过蛋白免疫印迹实验检测各转染组细胞活化前后胞核与胞质蛋白中p65的量以及胞质蛋白中iκb的量,反应nf-κb核转位水平。结果如图5所示。图中所示对照组为不加刺激的对照组,tnf-α组为实验组。cyt代表胞浆蛋白样品,nuc代表胞核蛋白样品,h3指组氨酸-h3。由图可见,胞浆内iκb在细胞活化后降解。同时,经tnf-α活化后,可见p65在胞质蛋白内总量下降,在胞核蛋白内总量上升,说明p65蛋白由胞质转移到了胞核。三组之间iκb降解程度以及p65核转位水平无明显差异,说明fkbp5对nf-κb核转位水平并无明显调控作用,同时fkbp5v55l突变不影响fkbp5对nf-κb核转位的调控作用。可以看出,经tnf-α活化后,野生型组与突变型组hek293细胞中,p65核转位水平及iκb降解水平无明显差异(图5),说明该突变不影响nf-κb核转位水平。4、aktser473磷酸化水平检测将野生型pcdna3.1-fkbp5质粒、突变型pcdna3.1-fkbp5质粒、pcdna3.1质粒分别转染到hek293细胞内。转染后的细胞血清饥饿24小时,使用脂多糖(lps)刺激活化。提取活化后细胞的总蛋白,经蛋白免疫印迹实验检测细胞内aktser473位点磷酸化水平,研究野生型及突变型fkbp5对akt磷酸化的调节作用有无差异。结果显示,野生型组或突变型组hek293细胞,其fkbp5表达水平明显上升,证明质粒转染成功,目的蛋白质fkbp5过表达(图6.a)。野生型组hek293细胞在经过lps活化后,其aktser473位点磷酸化水平低于空载体对照组,p=0.0412,说明fkbp5过表达后降低aktser473位点磷酸化水平;野生型组与突变型组相比,野生型组aktser473位点磷酸化水平低于突变型组,p=0.0075,证实v55l突变体fkbp5蛋白增高aktser473位点磷酸化(图6.b、c)。实施例4:fkbp5基因突变对小鼠破骨细胞影响及表型研究1、fkbp5c.163g>c点突变c57bl/6小鼠构建及鉴定点突变小鼠构建策略:a.es打靶载体设计和构建。由赛业生物科技有限公司构建含有引入了c.163g>c点突变的同源臂序列、loxp位点以及新霉素筛选标签(neocassette)的es打靶载体。b.打靶载体电转es细胞、筛选阳性克隆。将构建好的打靶载体电转至c57bl/6es细胞(赛业生物科技有限公司)内,利用新霉素抗性筛选阳性细胞并通过pcr扩增产物琼脂糖凝胶电泳、sanger测序以及southernblot等鉴定阳性电转克隆。c.es显微注射。将阳性es细胞显微注射到囊胚中,然后将显微注射后的囊胚回送到代孕鼠(c57bl/6小鼠,来源于赛业生物科技有限公司,鼠龄9-10周)输卵管或者子宫中,最终获得嵌合体小鼠。d.f1代小鼠繁殖。将得到的嵌合体小鼠和cre小鼠(来自赛业生物科技有限公司,鼠龄8-9周)进行交配繁殖,并进行基因型鉴定,其中带有目的基因点突变且已成功敲除新霉素抗性的f1代杂合子小鼠即为实验所需fkbp5v55l点突变杂合子小鼠。小鼠打靶及构建策略见图7所示。对f1代杂合子小鼠后代进行分析。小鼠基因型鉴定:通过sanger测序和pcr产物琼脂糖凝胶电泳鉴定小鼠基因型。sanger测序法引物序列:上游:5’-cctcgaggcatgcgataatctagct-3’下游:5’-ggcaaatggcttctttctgtcatg-3’测序方法同前文所述人fkbp5基因测序方法。测序鉴定的图谱如图14所示。pcr产物琼脂糖凝胶电泳法引物序列:上游:5’-tgtgtacctttgcctgctcatga-3’下游:5’-ccagagcaaggcaaaaggatcact-3’如图15所示,使用该引物扩增的pcr产物,不同基因型小鼠所得产物长度不同。野生鼠:195bp;杂合鼠:195bp、320bp;纯合鼠:320bp。因此,根据pcr产物琼脂糖凝胶电泳结果可以判定小鼠基因型。如图所示,第13/14/19泳道样本来自野生小鼠,第2/3/5/6/7/8/9/12/17/18/20/22泳道样本来自杂合小鼠,而第4/10/11/15/16/21泳道样本来自纯合突变鼠。根据多重鉴定结果,获得杂合鼠和纯合鼠,对这些小鼠进行后续试验。2、小鼠破骨细胞分化及功能研究分离培养fkbp5v55l小鼠及同窝fkbp5wt野生鼠骨髓来源的单核/巨噬细胞系细胞(bonemarrow-derivedmonocyte/macrophagecells,bmm):脱颈处死7-8周大小实验小鼠,游离其股骨和胫骨。剪去股骨和胫骨两端软骨,露出骨髓腔。使用2ml无菌注射器将骨髓细胞吹至一个15ml离心管内,离心收集骨髓细胞。向骨髓细胞沉淀中加入1mlα-mem(invitrogen)培养基重悬细胞,并加入3倍体积红细胞裂解液裂解红细胞。离心,收集细胞。使用α-mem完全培养基(含10%fbs)将细胞密度调整为1×107/10ml并接种到100mm平皿内,每皿接种10ml细胞悬液。培养基中加入终浓度为5ng/ml的m-csf因子,接种好的细胞置于细胞培养箱中培养。12-16小时后,收集平皿内未贴壁的悬浮细胞。收集得到的悬浮细胞用α-mem完全培养基重新悬起并计数,按照1.5×105个/孔的细胞密度接种到48孔板内,每孔接种体积为0.25ml。培养基中加入终浓度为30ng/ml的m-csf因子,继续培养3d。换液,弃掉悬浮细胞,所得贴壁细胞即为小鼠骨髓来源单核/巨噬细胞(bmm)。小鼠bmm诱导破骨细胞分化及trap染色鉴定:48孔板中单核/巨噬细胞继续加m-csf及rankl因子诱导分化。每孔加入含有m-csf(30ng/ml)以及不同浓度的rankl(0、30ng/ml、70ng/ml、150ng/ml)因子的α-mem完全培养基250μl,培养6-7天。使用抗酒石酸酸性磷酸酶染色试剂盒(货号:387a,sigma-aldrich,st.louis,mo,usa)进行trap染色。操作步骤严格按照试剂盒所附说明书进行。破骨细胞鉴定方法:胞核≥3个,trap染色阳性的多核巨细胞为破骨细胞。trap染色结果显示,当rankl诱导浓度较低(30ng/ml),野生鼠和纯合突变鼠骨髓来源单核巨噬细胞分化为破骨细胞的数量较少,且两组之间无显著差异(野生组vs突变组:17.0±4.2vs33.5±14.8;p=0.3468);而当rankl诱导浓度增高(70ng/ml、150ng/ml)后,两组破骨细胞数目均明显上升,且突变组所形成的破骨细胞数目显著多于野生组(70ng/ml,野生组vs突变组:42.7±7.6vs72.2±13.2;p=0.0284;150ng/ml,野生组vs突变组:55.7±22.1vs130.3±23.5;p=0.0162)。trap染色结果图及破骨细胞数目统计分别见图8、9。陷窝吸收实验:牛骨切片购自贵州医科大学,规格为50μm厚、60*40mm大小正方形骨片。牛骨切片保存于含有100iu双抗的pbs缓冲液中,4℃保存。切片在使用前需用超声清洗三次。取实验所需的牛骨切片,置于pbs缓冲液中,100hz/min频率超声三次,每次5分钟。超声清洗后的切片用75%酒精冲洗一遍,然后浸泡在75%酒精内大于2小时。将切片取出,置于48孔板内,开盖照紫外灭菌,切片每一面照紫外1小时。种板前,将灭菌后的骨片置于48孔板内,加入α-mem培养基,放到37℃培养箱内2小时,饱和切片。分离培养fkbp5v55l小鼠及同窝fkbp5wt野生鼠bmm,并接种在牛骨切片上。加入含有rankl(100ng/ml)、m-csf(50ng/ml)的α-mem完全培养基,诱导细胞分化。每隔两天换液,培养6-7天后进行甲苯胺蓝染色。染色步骤如下:将骨片置于2.5%戊二醛固定液固定7min,然后用0.25m氨水溶液超声清洗骨片,超声频率100hz/min,每次5min,共清洗3次。清洗后的切片经系列乙醇脱水(70%乙醇,5min;75%乙醇,5min;80%乙醇,5min;90%乙醇,5min;95%乙醇,5min;100%乙醇,5min;100%乙醇,10min),自然晾干。1%甲苯胺蓝染液室温染色3-4min,蒸馏水清洗后光镜下观察骨片陷窝吸收情况并拍照。使用imageproplus软件,计算陷窝吸收面积。统计10倍物镜下每视野内陷窝吸收面积。如图10所示,牛骨切片骨陷窝形成实验证实,每低倍镜视野下(40×)突变鼠破骨细胞形成的吸收陷窝面积大于野生鼠破骨细胞形成的陷窝面积(野生组vs突变组:4.73±1.02vs8.29±0.66;p=0.0071;图10)。说明在体外实验中,fkbp5v55l点突变小鼠破骨细胞骨吸收活性高于野生组。3、小鼠表型研究取10个月大fkbp5v55l小鼠及同窝fkbp5wt野生鼠股骨及胫骨并固定。固定后的标本使用西门子inveonmicro-ct检测仪扫描。扫描结果使用仪器自带inveonacquisitionworkplace和inveonresearchworkplace平台进行成像及骨吸收参数分析。小鼠股骨micro-ct检测骨小梁参数分析结果显示,突变组小鼠股骨骨小梁吸收模式增强。分析参数包括:相对骨体积(bonevolum/totalvolum,bv/tv)、骨小梁数量(trabecularnumber,tb.n)、骨小梁厚度(trabecularthinckness,tb.th)、骨小梁分离度(trabecularspacing,tb.sp)、骨小梁模式因子(trabecularbonepatternfactor,tbpf)。其中突变组小鼠bv/tv、tb.n以及tb.th减少(野生鼠vs突变鼠:bv/tv:0.1401±0.0277vs0.08±0.0219,p=0.0037;tb.n:3.4393±0.5322vs2.2073±0.3871,单位1/mm,p=0.0020;tb.th:0.0406±0.0024vs0.0361±0.0036,单位mm,p=0.0359),同时tb.sp和tbpf增加(野生鼠vs突变鼠:tb.sp:0.2561±0.0479vs0.4279±0.0815,单位mm,p=0.0018;tbpf:20.60±1.50vs25.50±2.61,单位1/mm,p=0.0035),见图11。说明突变组小鼠股骨骨小梁吸收模式增强,该结果与破骨细胞体外分化及吸收能力检测结果相符。然而,突变组与野生组小鼠之间股骨皮质厚度无显著差异,见图11。股骨2d平面图显示,与野生小鼠相比较,突变小鼠的骨小梁较细、骨小梁之间空隙较多,与上述参数分析结果相呼应(图12)。股骨皮质内侧未见明显的吸收陷窝。股骨远端与胫骨连接处3d重构图显示,部分突变小鼠股骨远端外侧骨皮质出现疑似吸收陷窝(箭头所示),野生组小鼠未见(图13)。以上结果再次反映fkbp5突变小鼠骨吸收模式增强,fkbp5突变小鼠出现疑似paget样表型。结论:paget骨病是一种以局部骨组织异常增生为特点的慢性骨代谢性疾病。其主要的病理改变为破骨细胞异常,包括病变部位破骨细胞数量增多,破骨细胞体积增大,细胞核数量增多,患者外周血单核细胞破骨细胞分化能力增强等。paget骨病致病原因包括遗传因素和环境因素,其中遗传致病因素在paget骨病发生中具有重要的作用。本发明中,通过对一个汉族paget骨病家系进行基因分析,确认了一个新的位于fkbp5基因的点突变(c.163g>c)与paget骨病发病相关。通过测序分析,明确了该突变为新发现的单核苷酸突变,而不是单核苷酸多态性。同时突变功能预测分析显示该突变极有可能影响fkbp5蛋白的功能。细胞实验证实该突变显著影响fkbp5对akt信号通路的调控作用,增加akt第473位丝氨酸磷酸化水平。动物实验证实fkbp5v55l突变能够增加小鼠bmm体外破骨细胞分化能力以及破骨细胞骨吸收能力。同时fkbp5v55l突变增加小鼠体内破骨细胞活性,使之小梁骨吸收模式增加,并呈现paget样骨病表型。综上,本发明有力地证实了fkbp5v55l突变通过增加破骨细胞分化及骨吸收能力,参与paget骨病发病。因此,本发明发现的fkbp5基因突变称为检测paget骨病的一个新的标志,为疾病的提前诊断和治疗提供依据。附:sqstm1、tnfrsf11a、tnfrsf11b、optn、vcp、csf1以及dcstamp基因外显子序列pcr扩增及测序用引物序列见下列表格。sqstm1引物:外显子引物序列(5’-3’)外显子11-fgcaatgaagagaggggtcag1-rttggtcaccactccagtcac外显子22-fgagcactcaccttccaggag2-rcaaattgctgaccccttcat外显子3-43-4-faaacagacacagggactgggaac3-4-rcctgacaaccatcacaaaagagaac外显子55-fgatgtccgggttaaaggtca5-rggcaacaaatcctcaccagt外显子66-fgtagtctccacaggccaagc6-ratgcacacgtgcagttaagg外显子77-fttaaagtcacgctgggaacc7-rccttctgtttgtgtcgctga外显子88-ftacagggaaagcaggtccac8-rctgccctaaatggcttcttgtnfrsf11a引物:外显子引物序列(5’-3’)外显子11-fgcggagaccggaccaagga1-ragtttccctccgcttgccct外显子22-facctagttttatccagaaagag2-rgagtgacagacattaccacctg外显子33-fgggtgcaatttttgttgctg3-racagaggccttggggatctaa外显子44-fgaagtgcgaggaggaacttg4-rtgaagaacctaagccgagga外显子55-fggttccataactcactgctggggtc5-rgaagttcctgtcacacatagggctg外显子66-fgattcaaatgtccaagaaggcg6-racattattagttacccccaatccag外显子77-fggtggggactgtcacttctgcta7-rgcttacaagtcatccgtttcctg外显子88-fgggactgtagttaatcatgttg8-rtcaatatgtgactaggatggc外显子99-faaagttccatagtcagttttccatc9-ragcttcctccttcctaaatggt外显子1010-faaccttcctctcggcagaccct10-rggattgcttgagcccaggagttatnfrsf11b引物:外显子引物序列(5’-3’)外显子11-ftttttttcccctgctctcccag1-rccgggcacccgtcggctggcccagg外显子22-fccactgtgttcctttctcctt2-rgctgcacattgacacgtacc外显子33-fgttaagagggcatctgctgg3-ratgatactacaaaatcgtac外显子44-fatccaattgttgagtaaatc4-raactaaacatacatgcagtct外显子55-fgatgtgaacacttatctggg5-raataatgtcatttatatagtoptn引物:vcp引物:csf1引物:dcstamp引物:外显子引物序列(5’-3’)外显子11-ftcatttcaacagctctcttcatctt1-rtcttttctcttttgccctcactc外显子22-fagaggttgattcatggtgatgtg2-ragggagactgtcggggtataggg外显子33-fttttgtagtcagtctaccaccat3-rgataaaccatgagaagtacctgt外显子44-faaagaagggatagagtaaaaaaa4-ratcagcattcaaatcacagagtt外显子55-fgaagaccaacataacccacaaaa5-rgagcagggagaaaagagaccatt参考文献:1.tiegs,r.d.(1997)paget'sdiseaseofbone:indicationsfortreatmentandgoalsoftherapy.clinther19,1309-1329;discussion1523-13042.vanstaa,t.p.,selby,p.,leufkens,h.g.,lyles,k.,sprafka,j.m.,andcooper,c.(2002)incidenceandnaturalhistoryofpaget'sdiseaseofboneinenglandandwales.jboneminerres17,465-4713.albagha,o.m.,visconti,m.r.,alonso,n.,langston,a.l.,cundy,t.,dargie,r.,dunlop,m.g.,fraser,w.d.,hooper,m.j.,isaia,g.,nicholson,g.c.,delpinomontes,j.,gonzalez-sarmiento,r.,distefano,m.,tenesa,a.,walsh,j.p.,andralston,s.h.(2010)genome-wideassociationstudyidentifiesvariantsatcsf1,optnandtnfrsf11aasgeneticriskfactorsforpaget'sdiseaseofbone.natgenet42,520-5244.wuyts,w.,vanwesenbeeck,l.,morales-piga,a.,ralston,s.,hocking,l.,vanhoenacker,f.,westhovens,r.,verbruggen,l.,anderson,d.,hughes,a.,andvanhul,w.(2001)evaluationoftheroleofrankandopggenesinpaget'sdiseaseofbone.bone28,104-1075.chung,p.y.,beyens,g.,defreitas,f.,boonen,s.,geusens,p.,vanhoenacker,f.,verbruggen,l.,vanoffel,j.,goemaere,s.,zmierczak,h.g.,westhovens,r.,devogelaer,j.p.,andvanhul,w.(2011)indicationsforageneticassociationofavcppolymorphismwiththepathogenesisofsporadicpaget'sdiseaseofbone,butnotfortnfsf11(rankl)andil-6polymorphisms.molgenetmetab103,287-2926.chung,p.y.,beyens,g.,boonen,s.,papapoulos,s.,geusens,p.,karperien,m.,vanhoenacker,f.,verbruggen,l.,fransen,e.,vanoffel,j.,goemaere,s.,zmierczak,h.g.,westhovens,r.,devogelaer,j.p.,andvanhul,w.(2010)themajorityofthegeneticriskforpaget'sdiseaseofboneisexplainedbygeneticvariantsclosetothecsf1,optn,tm7sf4,andtnfrsf11agenes.humgenet128,615-6267.chung,p.y.,andvanhul,w.(2012)paget'sdiseaseofbone:evidenceforcomplexpathogeneticinteractions.seminarthritisrheum41,619-6418.ralston,s.h.(2008)pathogenesisofpaget'sdiseaseofbone.bone43,819-8259.neale,s.d.,smith,r.,wass,j.a.,andathanasou,n.a.(2000)osteoclastdifferentiationfromcirculatingmononuclearprecursorsinpaget'sdiseaseishypersensitiveto1,25-dihydroxyvitamind(3)andrankl.bone27,409-41610.nakashima,t.,hayashi,m.,andtakayanagi,h.(2012)newinsightsintoosteoclastogenicsignalingmechanisms.trendsendocrinolmetab23,582-59011.li,l.,lou,z.,andwang,l.(2011)theroleoffkbp5incanceraetiologyandchemoresistance.brjcancer104,19-2312.pei,h.,li,l.,fridley,b.l.,jenkins,g.d.,kalari,k.r.,lingle,w.,petersen,g.,lou,z.,andwang,l.(2009)fkbp51affectscancercellresponsetochemotherapybynegativelyregulatingakt.cancercell16,259-266seqidno.1:(cds序列)atgactactgatgaaggtgccaagaacaatgaagaaagccccacagccactgttgctgagcagggagaggatattacctccaaaaaagacaggggagtattaaagattgtcaaaagagtggggaatggtgaggaaacgccgatgattggagacaaagtttatctccattacaaaggaaaattgtcaaatggaaagaagtttgattccagtcatgatagaaatgaaccatttgtctttagtcttggcaaaggccaagtcatcaaggcatgggacattggggtggctaccatgaagaaaggagagatatgccatttactgtgcaaaccagaatatgcatatggctcggctggcagtctccctaaaattccctcgaatgcaactctcttttttgagattgagctccttgatttcaaaggagaggatttatttgaagatggaggcattatccggagaaccaaacggaaaggagagggatattcaaatccaaacgaaggagcaacagtagaaatccacctggaaggccgctgtggtggaaggatgtttgactgcagagatgtggcattcactgtgggcgaaggagaagaccacgacattccaattggaattgacaaagctctggagaaaatgcagcgggaagaacaatgtattttatatcttggaccaagatatggttttggagaggcagggaagcctaaatttggcattgaacctaatgctgagcttatatatgaagttacacttaagagcttcgaaaaggccaaagaatcctgggagatggataccaaagaaaaattggagcaggctgccattgtcaaagagaagggaaccgtatacttcaagggaggcaaatacatgcaggcggtgattcagtatgggaagatagtgtcctggttagagatggaatatggtttatcagaaaaggaatcgaaagcttctgaatcatttctccttgctgcctttctgaacctggccatgtgctacctgaagcttagagaatacaccaaagctgttgaatgctgtgacaaggcccttggactggacagtgccaatgagaaaggcttgtataggaggggtgaagcccagctgctcatgaacgagtttgagtcagccaagggtgactttgagaaagtgctggaagtaaacccccagaataaggctgcaagactgcagatctccatgtgccagaaaaaggccaaggagcacaacgagcgggaccgcaggatatacgccaacatgttcaagaagtttgcagagcaggatgccaaggaagaggccaataaagcaatgggcaagaagacttcagaaggggtcactaatgaaaaaggaacagacagtcaagcaatggaagaagagaaacctgagggccacgtatgaseqidno.2:(mrna序列)agaacaaactcctacctctgtttgcaactaagagttacatcccccatgccaagaccttccgaggcgtcccagagggggaaattcctcgctgccggactgcgcccacccttgaggacctcacattctctttgggcagagcggaagccagacagagacagggactacgaagcgacaatccccagccctgtgtgtgcaaggagcccccgaggggcaggcagccacgctgcagaaggctagactctccgacccccagggagcctggtgtgggctgcagggacagaacgcttctcagaggaggttctctacttaaaagacaatgactactgatgaaggtgccaagaacaatgaagaaagccccacagccactgttgctgagcagggagaggatattacctccaaaaaagacaggggagtattaaagattgtcaaaagagtggggaatggtgaggaaacgccgatgattggagacaaagtttatctccattacaaaggaaaattgtcaaatggaaagaagtttgattccagtcatgatagaaatgaaccatttgtctttagtcttggcaaaggccaagtcatcaaggcatgggacattggggtggctaccatgaagaaaggagagatatgccatttactgtgcaaaccagaatatgcatatggctcggctggcagtctccctaaaattccctcgaatgcaactctcttttttgagattgagctccttgatttcaaaggagaggatttatttgaagatggaggcattatccggagaaccaaacggaaaggagagggatattcaaatccaaacgaaggagcaacagtagaaatccacctggaaggccgctgtggtggaaggatgtttgactgcagagatgtggcattcactgtgggcgaaggagaagaccacgacattccaattggaattgacaaagctctggagaaaatgcagcgggaagaacaatgtattttatatcttggaccaagatatggttttggagaggcagggaagcctaaatttggcattgaacctaatgctgagcttatatatgaagttacacttaagagcttcgaaaaggccaaagaatcctgggagatggataccaaagaaaaattggagcaggctgccattgtcaaagagaagggaaccgtatacttcaagggaggcaaatacatgcaggcggtgattcagtatgggaagatagtgtcctggttagagatggaatatggtttatcagaaaaggaatcgaaagcttctgaatcatttctccttgctgcctttctgaacctggccatgtgctacctgaagcttagagaatacaccaaagctgttgaatgctgtgacaaggcccttggactggacagtgccaatgagaaaggcttgtataggaggggtgaagcccagctgctcatgaacgagtttgagtcagccaagggtgactttgagaaagtgctggaagtaaacccccagaataaggctgcaagactgcagatctccatgtgccagaaaaaggccaaggagcacaacgagcgggaccgcaggatatacgccaacatgttcaagaagtttgcagagcaggatgccaaggaagaggccaataaagcaatgggcaagaagacttcagaaggggtcactaatgaaaaaggaacagacagtcaagcaatggaagaagagaaacctgagggccacgtatgacgccacgccaaggagggaagagtcccagtgaactcggcccctcctcaatgggctttcccccaactcaggacagaacagtgtttaatgtaaagtttgttatagtctatgtgattctggaagcaaatggcaaaaccagtagcttcccaaaaacagcccccctgctgctgcccggagggttcactgaggggtggcacgggaccactccaggtggaacaaacagaaatgactgtggtgtggagggagtgagccagcagcttaagtccagctcatttcagtttctatcaaccttcaagtatccaattcagggtccctggagatcatcctaacaatgtggggctgttaggttttacctttgaactttcatagcactgcagaaacctttaaaaaaaaaatgcttcatgaatttctcctttcctacagttgggtagggtaggggaaggaggataagcttttgttttttaaatgactgaagtgctataaatgtagtctgttgcatttttaaccaacagaacccacagtagaggggtctcatgtctccccagttccacagcagtgtcacagacgtgaaagccagaacctcagaggccacttgcttgctgacttagcctcctcccaaagtccccctcctcagccagcctccttgtgagagtggctttctaccacacacagcctgtccctgggggagtaattctgtcattcctaaaacacccttcagcaatgataatgagcagatgagagtttctggattagcttttcctattttcgatgaagttctgagatactgaaatgtgaaaagagcaatcagaattgtgctttttctcccctcctctattccttttagggaataatattcaatacacagtacttcctcccagcattgctactgctcagcttcttctttcattctaatccttgctattaagaatttaagacttgtgcttacaatatttttgacctggagtggatctatttacatagtcatttaggatccatgcagctttttttgtctttttaagattattggctcataagcatatgtatactggtttatggaactttatttacactcctctatcatgcaaaaaaattttgactttttagtactaagcttaatttttaaaaacaaaatctgtagggttgacaaataaatagttgctcttctacactaggggtttcacctgcaggtttgacacgcagttgctcgcttttcctgccctgtcaagcttctctgttctggcgtgagttgtgaaagagttgaagacagcttcccatgccggtacacagccagtagcctaaatctccagtacttgagctgaccattgaactagggcaagtcttaaatgtgtacatgtagttgaatttcagtccttacgggtaaacagattgagcatggctctctattccctcagcctaagaaacactcatgggaatgcatttggcaacccaaggaaccatttgcttaaacctggaacatctcacctttttaaatcctaaaaaacactggcagttatattttaaattagtttttatttttatgatggttttatcaaaagacttttattattagattgggacccccttcaaacctaaaaatcaagttatttccttttataatacttttcttccccatggaacaaatgggatcaatttgtgagttttttcctttaatgataactaaaatccctctaatttctcatttatgcttttgtcttttttatgaaatatttcttttaaaagccccagtctcacctacgaaatatgaagagcaaaagctgattttgcttacttgctaaactgttgggaaagctctgtagagcatggttccagtgaggccaagattgaaatttgatactaaaaaggccacctagctttttgcagataacaaacaagaaagctattccaagactcagatgatgccagctgtctcccacgtgtgtattatggttcaccagggggaactggcaaaagtgtgtgtggggaggggaagggtgtgtgagtggttctgagcaaataactacagggtgcccattaccactcaagaagacacttcacgtattcttgtatcaaattcaataatcttaaacaatttgtgtagaagtccacagacatctttcaaccaccttttaggctgcatatggattgccaagtcagcatatgaggaattaaagacattgtttttaaaaaaaaaaaatcatttagatgcacttttttgtgtgttctttaaataaatccaaaaaaaatgtgacttccaaaaaaaaaaaseqidno:3:(基因组序列)aaaataaatacactggattttacagtagttaaatactttactagaaaacaagttattcagtatcatatcaagcagtgcaatgatactcatgttttttacagatagactatgtttcaaataataggaaatgtaacctgttttaccaggaaattgggctttctgaaaataaacaagcagaatctagtcaggagctaatgtttttagcaaaggagggaaataaaattacacctttgaagctttgcttaagcttctttttgaatattttaaaagtccaccattctttctggtctaagtccgtgtgtatatggactagctcacaagcctgtgttagagagtttagtgtcgggcgcggtggctcatgcttgtaatcccagcactttgggaggctgaggcgggtggatcacgaggtcaggagttcgagaccagcctggccaacatggtgaaaccccatctctactaaaaatacaaaaattagctgagcacggaggcgcacgcctgtaatcccagctactcgggaggctgaggcaggagaattgcttcaacctgggaggcagaggttgcagtgagccgagatcgtgccattgcactccagcctgggcgacagggcaagactccgtctcaaaaaaaaaaaaaaaaaaaaaaaagagtttactgtcaaaatatagggcattaaaccaatgaacaggatttattggttaattctttgaaggaataaatagtgcgattagcaataaaagtagatataacataaaagacaagttggtaaaggacagttttctaacaatgagttattgtgaggaatgaagggaggcattagaaagaatggataatagtatatttgcatatctgcctggatgtcttgggatggactgtgtgacctttttattctctgattacaagtaaaaagtttgacctcctttttcttaatgtttgtcttttctagattgtcaaaagagtggggaatggtgaggaaacgccgatgattggagacaaagtttatctccattacaaaggaaaattgtcaaatggaaagaagtttgattccagtcatgatagaaatgaaccatttgtctttagtcttggcaaaggtaagagggtatttttttgagttgtgttttacttagctttattactatttgtaatatataggagcagaggattctatcctttctaaaagtaaagaaattttagagtagtagtactcaaagaatggtctgagaatccctagggctctctgagatctttttagggggtcccaaatgtcatttattttcataacaatactaagatattatttgccattttactgtattgacatttatttgcactgatggtacaaaagctgcagttgtaaaaaaactgctggttccttaacacaaatcaaggcagtggtaaccagctgttccagtagtcatggtattttccactgccactccttcactatttacataagggaaaaagctagtttcacgtaaaaatgtcctgataaagcagtaaaaattattagttttattgaaacccaatctttgagtttcacatcactttgatattctgtgacagaaagggaagcacgtgtagagcacttctgtcccccaaagtatgatggttgtctcaaggaaaagtacttgtgctcttgttttagttgagatctgaactaactgcatttatttcatgagataccatttttacttgaaagggcaaatctgattatgcagacttgattcagtatttggcaggtgtttttctgagaatgaatggagtaagtctgtcacttcaaggaaaaacaactgacaatgttttttgccattaagttttcaagtaaaaattatagtttaggagaactcctatttgccactgtgggcttatttgccactgtgggcttgacagcttccgaagacttttctgatgagattggtgggtattaatgaatgtggttttaaatatacaatgaaatgtgtccacatttggaagatctgtataactaagaatgaatattttctcseqidno.4mttdegaknneesptatvaeqgeditskkdrgvlkivkrvgngeetpmigdkvylhykgklsngkkfdsshdrnepfvfslgkgqvikawdigvatmkkgeichllckpeyaygsagslpkipsnatlffeielldfkgedlfedggiirrtkrkgegysnpnegatveihlegrcggrmfdcrdvaftvgegedhdipigidkalekmqreeqcilylgprygfgeagkpkfgiepnaeliyevtlksfekakeswemdtkekleqaaivkekgtvyfkggkymqaviqygkivswlemeyglsekeskasesfllaaflnlamcylklreytkaveccdkalgldsanekglyrrgeaqllmnefesakgdfekvlevnpqnkaarlqismcqkkakehnerdrriyanmfkkfaeqdakeeankamgkktsegvtnekgtdsqameeekpeghvseqidno.7attgtcaaaagagtggggaatggtgaggaaacgccgatgattggagacaaagtttatctccattacaaaggaaaattgtcaaatggaaagaagtttgattccagtcatgatagaaatgaaccatttgtctttagtcttggcaaaseqidno.8attaaaccaatgaacaggatttattggttaattctttgaaggaataaatagtgcgattagcaataaaagtagatataacataaaagacaagttggtaaaggacagttttctaacaatgagttattgtgaggaatgaagggaggcattagaaagaatggataatagtatatttgcatatctgcctggatgtcttgggatggactgtgtgacctttttattctctgattacaagtaaaaagtttgacctcctttttcttaatgtttgtcttttctagattgtcaaaagagtggggaatggtgaggaaacgccgatgattggagacaaagtttatctccattacaaaggaaaattgtcaaatggaaagaagtttgattccagtcatgatagaaatgaaccatttgtctttagtcttggcaaaggtaagagggtatttttttgagttgtg当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1