新颖的启动子的制作方法
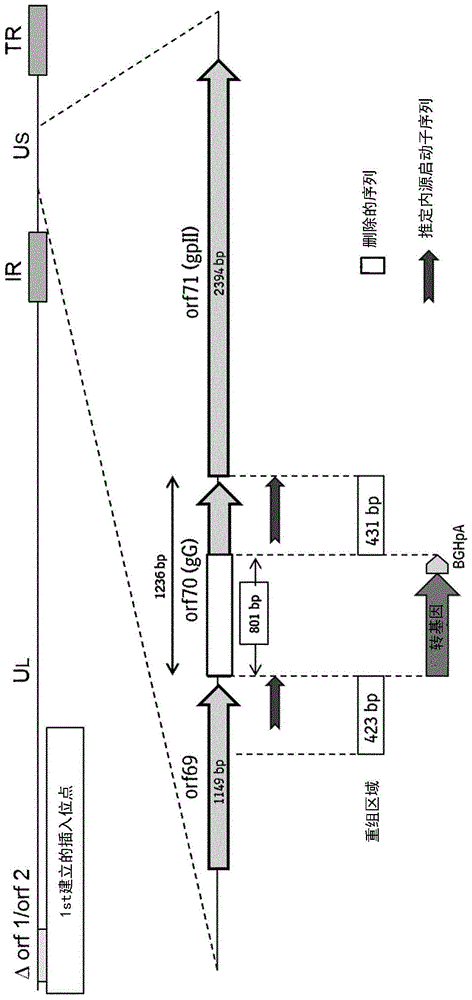
本申请案根据37c.f.r.1.821-1.825含有序列表。本申请案随附的序列表以全文引用的方式并入本文中。本发明关于(载体)疫苗领域,且尤其关于新颖的启动子序列、表达盒及载体,他们适于表达所关注的基因,尤其抗原编码序列。本发明的病毒载体可用于生产免疫原性组合物或疫苗。
背景技术:
:马病原体马α疱疹病毒1(马流产病毒,ehv-1)属于疱疹病毒目(herpesvirales)疱疹病毒科(herpesviridae)中α疱疹病毒亚科(alphaherpesvirinae)的水痘病毒属(varicellovirus)。其为具有约150,000个碱基对的双链dna基因组的大包膜病毒。水痘病毒亚属的其他重要成员为人类α疱疹病毒3(水痘带状疱疹病毒)、猪α疱疹病毒1(伪狂犬病病毒)、牛α疱疹病毒1(感染性支气管炎病毒)及马α疱疹病毒4(马鼻肺炎病毒,ehv-4)(http://www.ictvonline.org/virustaxonomy.aspvirustaxonomy:2015releaseec47,london,uk,2015年7月;emailratification2016(msl#30)ehv-1及ehv-4为地方病且侵袭全世界的马。虽然ehv-4引起大多轻度的上呼吸道感染,但ehv-1可引起伴有自呼吸系统症状至流产及致死性脑脊髓病的一系列疾病的全身性感染,这视病毒株及宿主的免疫状态而定。目前,在美国及欧洲分别可使用两种经许可的针对ehv-1的改良式活毒疫苗(mlv)tm(boehringeringelheim)及tm(msd)。两者均含有传统减毒的ehv-1rach病毒株,其在猪上皮细胞中传代256次以便减毒(ma等人2013)。减毒机制已在分子水平上加以研究。osterrieder等人(1996)展示rach不具有orf67的两个基因组拷贝且一个拷贝的恢复足以恢复毒力。另外,rach携带1283bp缺失,移除大于90%的编码免疫抑制性病毒蛋白的orf1编码序列。其他突变亦可影响减毒,但迄今为止尚未详细研究。所有这一切使得rach成为非常安全的疫苗株,因为(若可能)通过在经疫苗接种的动物体内传代回复毒力几乎不可能。含有马α疱疹病毒1(ehv-1)疫苗株rach(prach-se)的整个基因组的大肠杆菌细菌人工染色体(bac)称为用于载体疫苗开发的平台。已展示基于ehv-1rach的载体疫苗能够在包括猪、牛及犬的数个哺乳动物物种中引发免疫(rosas等人2007,rosas等人2008,trapp等人2005,said等人2013)。编码病原体的抗原蛋白的基因可由重组ehv-1rach表达。ehv-1-rach基因组是以其bac形式在大肠杆菌中进行操纵且经调整以通常通过插入转基因表达盒表达额外蛋白质(tischer等人,2010)。在prach-sedna转染于培养的容许细胞后,通过细胞转录因子开始ehv-1复制。病毒dna聚合酶的活性使得所有bac载体相关序列缺失及ehv-1rach基因组恢复至其原始状态。生成无法与rach区分的感染性病毒。当在大肠杆菌中例如通过插入转基因表达盒操纵prach-se时,在容许细胞中转染后重建的病毒将携带修饰且将表达额外基因。重组ehv-1rach可用作载体疫苗。然而,在无额外的外源启动子的情况下表达的转基因蛋白质的量通常相对较低。因此,对可用于自此类载体、尤其重组ehv-1rach表达转基因蛋白质的额外启动子的需要尚未满足。野生型ehv-1病毒株在其基因组的特有长区段(序列坐标1298-3614;图2)的一端具有称为orf1、orf2及orf3的三个开放阅读框架(orf)。orf1及orf3串联排列于dna的一链上,而orf2由互补链编码。疫苗株rach在影响orf1及orf2的区域具有1283bp缺失,表明这些基因对于病毒复制而言为非必需的。因此,该位点充当转基因插入位点。已报导,使用人类细胞巨大病毒立即早期基因1启动子-增强子(boshart等人1985)自orf1/3插入位点有效表达转基因。在此类研究中,牛生长激素聚腺苷酸化信号(bgh)用于稳定转录物以便优选表达(ma等人2012;said等人2013)。虽然不存在hcmv可诱发人类肿瘤的迹象,但无法排除理论风险。在描述hcmv-ie增强子(boshart等人1985)之前,在如猿猴病毒40、多瘤病毒或莫罗尼鼠肉瘤病毒(moloneymurinesarcomavirus)的已知致癌病毒的基因组中发现大部分强劲的增强子。虽然极其强劲的非组织特异性hcmv及mcmv(小鼠细胞巨大病毒)ie启动子-增强子非常适于多种研究活动,但其一般可能不代表转基因载体疫苗的启动子的第一选择。具体而言,给动物接种疫苗的人员意外暴露的风险可被监管当局视为授予疫苗许可证的障碍。技术实现要素:为了避免任何此类障碍,本发明提供新颖的用于转基因表达的调控核酸序列/启动子序列,尤其在载体疫苗的情形内及尤其在ehv-1载体的情形内。因此,上述技术问题的解决方案通过本说明书及在申请专利范围中表征的实施例来达成,且本发明在其不同方面中是根据申请专利范围来实施。本发明提供新颖的用于转基因表达的调控核酸序列/启动子序列、克服本领域的缺陷的免疫原性组合物、疫苗及相关方法。确定的广泛用于在包括疱疹病毒的不同载体系统中驱动高水平的转基因表达的启动子序列为hcmv(boshart等人1985;foecking及hofstetter1986)或小鼠细胞巨大病毒(mcmv;等人1985)的立即早期基因的启动子序列或如猿猴病毒40(sv40)的致癌病毒的强启动子,例如sv40大t抗原启动子及更多(例如kim等人1990)。此类强启动子为细胞生物学家偏好的,因为其自主地在不同细胞培养系统中起作用。在病毒复制的情形下,经感染细胞通过病毒功能转化成病毒复制机器。疱疹病毒复制及形态发生的生物学被充分理解。在感染后,仅极少基因(α-基因)经转录且翻译成立即早期蛋白质(iep)。这些iep为编码病毒酶(如dna聚合酶及许多其他酶)的β-基因的转录活化因子。病毒基因组复制的起始标志着转录编码病毒结构蛋白的β-及γ-基因的病毒复制后期的开始(fields,2013)。然而,对于改良式载体疫苗,上述自主性强启动子中无一者被视为选项,特别是来源于致癌病毒的启动子具有不利的安全概况。因此,需要提供在病毒复制的情形下具有高活性的启动子,如ehv-1β-及γ-基因的那些启动子。由于不可能在一个载体分子中使用一致的dna序列两次而无进行内部同源重组且因此基因不稳定的风险,故本发明提供新颖的来源于ehv-4的公开基因组序列(马α疱疹病毒4病毒株ns80567,完整基因组,登陆编号af030027,版本af030027.1gi:2605950,日期1998年5月21日)的替代性启动子序列。该等基因与ehv-1基因的序列相同性在55至84%范围内。本发明提供两种新颖的启动子:p430及p455,其在细胞培养物中以及在动物(猪及小鼠)体内rehv1-rach复制的背景下展示具有功能性。两种新颖的启动子在病毒复制周期期间的活性水平似乎与自活体外启动子动力学实验所推断的极其类似。这些特性允许形成以类似效率并行表达两种不同抗原的基于ehv-1rach的重组载体疫苗。若疫苗靶标由两种不同病原体组成,则两种新颖的启动子与两个聚腺苷酸化序列组合应用在两个插入位点中可显著降低货品的成本且呈现优于仅表达一种抗原成分的载体的明显优势。本发明提供两种新颖的启动子:4pgg600及4pmcp600及其较短长度的衍生物,其在细胞培养物中瞬时转染之后或在细胞培养物中rehv1-rach复制的背景下展示具有功能性。另外,由本发明提供的新颖的启动子序列亦在如犬腺病毒(cadv)的其他载体背景下展示为高效的。重要的是,当cag或cmv5启动子序列存在于位于e3区中的表达盒中时,未实现重组cadv的拯救。其似乎具有序列特异性,因为表达盒的尺寸尚未超过所观察到实验基因组尺寸限制。因此,本发明的新颖的源于ehv-4的启动子序列(诸如p430及p455)不仅有助于转基因表达,且亦支持病毒拯救的关键步骤,因此,鉴于先前技术启动子序列为有利的。具体实施方式本发明解决了先前技术中固有的问题且提供目前先进技术中的明显进步。一般而言,本发明提供启动子序列/调控核酸序列,其包含4pgg600(seqidno.1)或4pmcp600(seqidno.2)或其互补核苷酸序列,或其功能片段或功能衍生物或其互补核苷酸序列,其中该启动子序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达。在一特定方面中,功能片段或衍生物的长度为550个核苷酸,优选500、490、480、470、460、455、450、445、440、435、434、433、432、431、430个核苷酸,最优选455或430个核苷酸。在另一方面中,功能片段或衍生物的长度在430至550个核苷酸、430至500个核苷酸或430至480个核苷酸之间。优选地,功能片段具有70%、80%、85%,优选90%、91%、92%、93%、94%,更优选95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%的序列相同性和/或同源性或序列相同性。在一特定方面中,表达为增加的。在一特定方面中,功能片段为截短的4pgg600(seqidno.1)或其互补核苷酸序列,相对于整个长度的序列相同性优选为(至少)72%(或更高)。优选地,4pgg600(seqidno.1)的功能片段为命名为p430(seqidno:3)的片段。在另一方面中,序列相同性为(至少)70%、80%、85%,优选90%、91%、92%、93%、94%,更优选95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%。在另一特定方面中,功能片段为截短的4pmcp600(seqidno.2)或其互补核苷酸序列,相对于整个长度的序列相同性优选为(至少)78%(或更高)。优选地,4pmcp600(seqidno.2)的功能片段为命名为p455(seqidno:4)的片段。在另一方面中,序列相同性为(至少)70%、80%、85%,优选90%、91%、92%、93%、94%,更优选95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%。本发明另外提供启动子序列/调控核酸序列,包含p430(seqidno.3)或p455(seqidno.4)或其互补核苷酸序列,或其功能片段或功能衍生物或其互补核苷酸序列,其中该启动子序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达。优选地,功能片段具有70%、80%、85%,优选90%、91%、92%、93%、94%,更优选95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%的序列同源性或序列相同性。在一特定方面中,表达为增加的。本发明另外关于包含选自由以下组成的群的启动子序列/调控核酸序列的表达盒:4pgg600(seqidno.1)及4pmcp600(seqidno.2)及其互补核苷酸序列,及其功能片段及功能衍生物及其互补核苷酸序列,及p430(seqidno.3)及p455(seqidno.4)及其互补核苷酸序列,及其功能片段及其功能衍生物及其互补核苷酸序列,其中该启动子序列可操作地连接于所关注序列、优选所关注基因或抗原编码序列、更优选所关注异源和/或外源序列、所关注基因或抗原编码序列,其中该启动子序列/调控核酸序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达,由此该启动子序列优选为异源启动子序列/调控核酸序列、更优选外源启动子序列/调控核酸序列。在一特定方面中,表达为增加的。在一特定方面中,表达盒为重组、异源和/或外源表达盒。在另一特定方面中,启动子序列/调控核酸序列为重组、异源和/或外源启动子序列/调控核酸序列。本发明另外关于包含根据本发明的表达盒的载体,诸如病毒载体或病毒构建体。优选地,该载体可用于生产免疫原性组合物或疫苗。在另一方面中,本发明是关于包含具有选自由以下组成的群的启动子序列/调控核酸序列的表达盒的载体(诸如病毒载体或病毒构建体):4pgg600(seqidno.1)及4pmcp600(seqidno.2)及其互补核苷酸序列,及其功能片段及功能衍生物及其互补核苷酸序列,及p430(seqidno.3)及p455(seqidno.4)及其互补核苷酸序列,及其功能片段及其功能衍生物及其互补核苷酸序列,其中该启动子序列可操作地连接于所关注序列、优选所关注基因或抗原编码序列、更优选所关注异源和/或外源序列、所关注基因或抗原编码序列,其中该启动子序列/调控核酸序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达,由此该启动子序列优选为异源启动子序列/调控核酸序列、更优选外源启动子序列/调控核酸序列。在一特定方面中,表达为增加的。优选地,该载体可用于生产免疫原性组合物或疫苗。本发明另外关于异源(表达)载体诸如病毒载体或用于dna疫苗接种的质粒,包含含有4pgg600(seqidno.1)和/或4pmcp600(seqidno.2)或其互补核苷酸序列,或其功能片段或功能衍生物或其互补核苷酸序列,或p430(seqidno.3)和/或p455(seqidno.4)或其互补核苷酸序列,或其功能片段或其功能衍生物或其互补核苷酸序列的调控核酸/启动子序列,其中该调控核酸/启动子序列引起所关注序列、所关注基因或抗原编码序列的转录或表达。在一特定方面中,该所关注序列、所关注基因或抗原编码序列的转录或表达增加。优选地,该载体可用于生产免疫原性组合物或疫苗。在一特定方面中,载体为重组、异源和/或外源载体。在另一特定方面中,启动子序列/调控核酸序列为重组、异源和/或外源启动子序列/调控核酸序列。在根据本发明的表达盒和/或根据本发明的载体的一特定方面中,(启动子序列/调控核酸序列的)功能片段或衍生物具有70%、80%、85%,优选90%、91%、92%、93%、94%,更优选95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%的序列同源性或序列相同性。在根据本发明的表达盒和/或根据本发明的载体的另一特定方面中,(启动子序列/调控核酸序列的)功能片段或衍生物的长度为550个核苷酸,优选500、490、480、470、460、455、450、445、440、435、434、433、432、431、430个核苷酸,最优选455或430个核苷酸。在根据本发明的表达盒和/或根据本发明的载体的另一特定方面中,(启动子序列/调控核酸序列的)功能片段或衍生物的长度在430至550个核苷酸、430至500个核苷酸或430至480个核苷酸之间。在根据本发明的表达盒和/或根据本发明的载体的另一特定方面中,(启动子序列/调控核酸序列的)功能片段或衍生物具有70%、80%、85%,优选90%、91%、92%、93%、94%,更优选95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、99.9%的序列同源性或序列相同性。在根据本发明的表达盒和/或根据本发明的载体的另一特定方面中,(启动子序列/调控核酸序列的)功能片段或衍生物为截短的4pgg600(seqidno.1)或其互补核苷酸序列,相对于整个长度的序列相同性优选为(至少)72%(或更高)。优选地,4pgg600(seqidno.1)的该功能片段为命名为p430(seqidno:3)的片段。在根据本发明的表达盒和/或根据本发明的载体的另一特定方面中,(启动子序列/调控核酸序列的)功能片段或衍生物为截短的4pmcp600(seqidno.2)或其互补核苷酸序列,相对于整个长度的序列相同性优选为(至少)78%(或更高)。优选地,4pmcp600(seqidno.2)的该功能片段为命名为p455(seqidno:4)的片段。在根据本发明的表达盒和/或根据本发明的载体的另一特定方面中,其中该表达盒和/或该载体包含一或多个其他调控序列,诸如终止信号、聚腺苷酸化信号或如ires和/或2a肽的调控组件。在一特定方面中,根据本发明的载体为异源和/或外源载体。在本发明的另一特定方面中,根据本发明的载体为病毒载体,优选选自由以下组成的群:疱疹病毒科,诸如马α疱疹病毒1(ehv-1)、马α疱疹病毒4(ehv-4)及其他水痘病毒属,如猪α疱疹病毒1(伪狂犬病病毒,prv)及牛α疱疹病毒1(牛疱疹病毒1,bhv-1);腺病毒科(adenoviridae,adv),诸如cadv(犬腺病毒);腺相关病毒科;杆状病毒科(baculoviridae);慢病毒科(lentiviridae),诸如反转录病毒;及痘病毒科(poxviridae)。在一更特定方面中,该病毒载体为疱疹病毒科、优选α疱疹病毒属、更优选水痘病毒亚属的成员,该载体最优选为马α疱疹病毒1(ehv-1)。本发明另外关于一种产生载体、优选病毒载体的方法,其包含:a.提供根据本发明的启动子序列和/或调控核酸序列,b.将来自步骤a)的该启动子序列整合至来源于病毒的载体主链中,该病毒选自由以下组成的群:疱疹病毒科,诸如ehv-1、ehv-4、水痘病毒属,如猪α疱疹病毒1(伪狂犬病病毒,prv)及牛α疱疹病毒1(牛疱疹病毒1,bhv-1);腺病毒科(adv),诸如cadv(犬腺病毒);细小病毒科(parvoviridae),如腺相关病毒;杆状病毒科;反转录病毒科(retroviridae)及痘病毒科,该载体主链优选来源于疱疹病毒,该载体主链更优选为ehv-1或ehv-4。本发明另外关于一种真核生物宿主细胞株,其特征在于容许根据本发明的载体复制,该宿主细胞株优选为哺乳动物细胞株或昆虫细胞株,其最优选为pk/wrl细胞株、rk13细胞株、mdbk细胞株、st细胞株、ai-st细胞株、vero细胞株、sf9细胞株、sf21、sf+细胞株、mdck细胞株和/或其衍生物。本发明另外关于一种制备宿主细胞的方法,其通过以下步骤表征:a.用根据技术方案9至19的载体感染根据技术方案21的真核生物宿主细胞株,b.在适合的条件下培养经感染细胞,c.可选地收集该宿主细胞。如上文所列的哺乳动物宿主细胞株一般在浸没于用于哺乳动物细胞培养的培养基,诸如补充有厄尔氏盐(earle'ssalt)及胎牛血清的最低必需培养基(mem)中的塑料组织培养器皿中培养。哺乳动物细胞株在培育箱中保持在37℃下补充有5%co2及约80%湿度的常规氛围中。昆虫细胞株培养在浸没于昆虫细胞培养基的塑料组织培养器皿中且在培育箱中保持在27℃下的常规氛围中。本发明另外关于包含4pgg600(seqidno.1)或4pmcp600(seqidno.2)或其互补核苷酸序列的核酸序列,或其功能片段或功能衍生物或其互补核苷酸序列作为启动子序列的用途,其中该核酸序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达。在一特定方面中,表达为增加的。本发明另外关于包含4pgg600(seqidno.1)或4pmcp600(seqidno.2)或其互补核苷酸序列的核酸序列,或其功能片段或功能衍生物或其互补核苷酸序列作为调控核酸序列的用途,其中该核酸序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达。在一特定方面中,表达为增加的。本发明另外关于包含p430(seqidno.3)或p455(seqidno.4)或其互补核苷酸序列的核酸序列,或其功能片段或功能衍生物或其互补核苷酸序列作为启动子序列的用途,其中该核酸序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达。在一特定方面中,表达为增加的。本发明另外关于包含p430(seqidno.3)或p455(seqidno.4)或其互补核苷酸序列的核酸序列,或其功能片段或功能衍生物或其互补核苷酸序列作为调控核酸序列的用途,其中该核酸序列引起所关注核苷酸序列、优选所关注基因、更优选抗原编码序列的表达。在一特定方面中,表达为增加的。本发明另外关于一种试剂盒,其由根据本发明的载体、宿主细胞、可选的转染剂及说明书印刷品组成。本发明另外关于根据本发明的载体或根据本发明的真核生物宿主细胞株或根据本发明的真核生物宿主细胞的用途,其用于制造的免疫原性组合物或疫苗,可选地使用m.o.i.为0.01至0.001。尤其对于病毒载体的用途,哺乳动物宿主细胞株一般培养及生长至汇合或亚汇合,视细胞株而定。病毒载体与适量的新鲜培养基混合且稀释以产生0.001至0.01的感染复数(m.o.i.)。自宿主细胞移除细胞培养基且用含有经稀释的病毒载体的培养基更换。如此接种的细胞培养物在37℃/5%co2下培育约2至4天,视细胞株而定。病毒载体在细胞中复制引起细胞病变效应(cpe)且最终细胞破坏及死亡。采集材料且储存在-80℃下。测定病毒效价。本发明另外关于一种免疫原性组合物,其包含a.根据本发明的表达盒,和/或b.根据本发明的载体,和/或c.由根据本发明的表达盒表达的多肽,和/或由根据本发明的载体表达的多肽,诸如病毒、经修饰的活病毒、病毒样颗粒(vlp)或其类似物,及d.可选的医药学或兽医学可接受的载剂或赋形剂,该载剂优选适于经口、皮内、肌肉内或鼻内施用。在一特定方面中,该免疫原性组合物包含病毒。在另一特定方面中,该免疫原性组合物包含感染性病毒。本发明另外关于一种疫苗或医药组合物,其包含a.根据本发明的表达盒,和/或b.根据本发明的载体,和/或c.由根据本发明的表达盒表达的多肽,和/或由根据本发明的载体表达的多肽,诸如经修饰的活病毒、病毒样颗粒(vlp)或其类似物,及d.医药学或兽医学可接受的载剂或赋形剂,该载剂优选适于经口、皮内、肌肉内或鼻内施用,e.该疫苗可选地另外包含佐剂。本发明此外关于一种制备用以降低与感染相关或由感染引起的一或多种临床征象的发生率或严重程度的免疫原性组合物或疫苗的方法,其包含以下步骤:a.用根据本发明的载体感染根据本发明的真核生物宿主细胞株,b.在适合的条件下培养经感染细胞,c.收集经感染细胞和/或载体和/或病毒成分,d.可选地纯化步骤c)的收集物,e.使该收集物与医药学上可接受的载剂混合。治疗方法本发明另外关于根据本发明的免疫原性组合物或根据本发明的疫苗,其用于减少或预防由动物感染病原体引起的临床征象或疾病的方法或用于治疗或预防动物感染病原体的方法中,该动物优选为产食性动物,诸如猪或牛。本发明另外提供一种用于使动物、诸如产食性动物免疫的方法,其包含向此类动物施用如本文所述的免疫原性组合物或疫苗。本发明另外关于一种使动物、诸如包括猪的产食性动物免疫以对抗由该动物中病原体引起的临床疾病的方法,该方法包含向该动物施用根据本发明的免疫原性组合物或根据本发明的疫苗的步骤,其中该免疫原性组合物或疫苗不会引起感染的临床征象,但能够诱发使该动物免疫以对抗该病原体的致病性形式的免疫反应。在一特定方面中,免疫使得畜群中特定病毒感染的发生率降低或使得由特定病毒感染引起或与特定病毒感染相关的临床征象的严重程度减轻。另外,用在此提供的免疫原性组合物使有需要的产食性动物免疫,由此预防产食性动物受病毒感染。甚至更优选地,免疫产生对抗该病毒感染的持久有效的免疫反应。应了解,该时间段将持续大于2个月、优选大于3个月、更优选大于4个月、更优选大于5个月、更优选大于6个月。应了解,免疫可不在所有经免疫动物/个体中有效。然而,期限需要畜群的相当大一部分的动物/个体经有效免疫。本发明提供一种用于治疗或预防有需要的动物、诸如产食性动物的由病毒引起的临床征象的方法,该方法包含向该动物施用治疗有效量的如本文所述的免疫原性组合物或疫苗。优选地,与未经处理(未经免疫)但随后感染特定猪a型流感病毒的动物相比,临床征象减少至少50%、甚至更优选至少60%、再更优选至少70%、甚至更优选至少80%、甚至更优选至少90%、再更优选至少95%、最优选100%。本发明另外关于一种用于给动物、优选产食性动物、诸如猪或牛接种疫苗以对抗与动物中病原体相关的疾病和/或降低与动物中病原体相关或由动物中病原体引起的一或多种临床征象的发生率或严重程度的试剂盒,其包含:a)能够向该动物施用疫苗的分配器;及b)根据本发明的免疫原性组合物或根据本发明的疫苗,及c)可选的说明书印刷品。定义除非另外定义,否则在申请时本文所用的所有技术及科学术语均具有与熟习本发明所属的技术者通常所理解相同的含义。术语的含义及范畴应为清楚的;然而,在任何潜在不明确性的情况下,本文所提供的定义优先于任何字典或外部定义。此外,除非上下文另外需要,否则单数术语应包括复数且复数术语应包括单数。除非另外说明,否则本文中使用“或”意谓“和/或”。此外,术语“包括(including)”以及其他形式(诸如“包括(includes)”及“包括(included)”)的使用不具限制性。本文所提及的所有专利及公开案以引用的方式并入本文中。除非另外指示,否则本发明的实施将采用在此项技术内的病毒学、分子生物学、微生物学、重组dna技术、蛋白质化学及免疫学的常规技术。此类技术在文献中充分解释。参见例如sambrook,fritsch及maniatis,molecularcloning:alaboratorymanual,第i卷、第ii卷及第iii卷,第二版(1989);dnacloning,第i卷及第ii卷(d.n.glover编1985);oligonucleotidesynthesis(m.j.gait编1984);nucleicacidhybridization(b.d.hames及s.j.higgins编1984);animalcellculture(r.k.freshney编1986);immobilizedcellsandenzymes(irlpress,1986);perbal,b.,apracticalguidetomolecularcloning(1984);theseries,methodsinenzymology(s.colowick及n.kaplan编,academicpress,inc.);proteinpurificationmethods-apracticalapproach(e.l.v.harris及s.angal,编,irlpressatoxforduniversitypress);及handbookofexperimentalimmunology,第i-iv卷(d.m.weir及c.c.blackwell编,1986,blackwellscientificpublications)。在详细描述本发明之前,应了解本发明不限于特定dna,因此多肽序列或方法参数当然可变化。亦应了解,本文所用的术语仅用于描述本发明的具体实施例的目的,且不意欲为限制性的。必须注意的是,除非内容明确另外指示,否则如本说明书及所附申请专利范围中所用,单数形式“一(a/an)”及“该(the)”包括复数个指示物。因此,举例而言,提及“抗原”包括两种或更多种抗原的混合物,提及“赋形剂”包括两种或更多种赋形剂的混合物,及其类似物。分子生物学定义如本领域已知,术语“载体”是指用于传输遗传物质至宿主细胞的多核苷酸构建体,通常为质粒或细菌人工染色体。载体可为例如细菌、病毒、噬菌体、细菌人工染色体、粘粒或质粒。如本文所用,载体可由dna或rna构成或含有dna或rna。在一些实施例中,载体由dna构成。在一些实施例中,载体为感染性病毒。此类病毒载体含有经一定方式操纵以携带外源基因的病毒基因组,该外源基因在细胞培养物及宿主动物中均不在病毒载体复制方面起作用。根据本发明的特定方面,载体可用于各个方面,诸如仅仅传输遗传物质、用于转染宿主细胞或生物体、用作疫苗(例如dna疫苗)或用于基因表达目的。基因表达为描述细胞中由称为基因的特异性多核苷酸序列引导的蛋白质生物合成的术语。在一特定方面中,载体可为“表达载体”,其为当存在于适当环境中时能够引导由载体携带的一或多个基因所编码的蛋白质表达的载体。载体及用于制造和/或使用载体(或重组体)进行表达的方法可利用或类似于以下所揭示的方法:美国专利第4,603,112号、第4,769,330号、第5,174,993号、第5,505,941号、第5,338,683号、第5,494,807号、第4,722,848号、第5,942,235号、第5,364,773号、第5,762,938号、第5,770,212号、第5,942,235号、第382,425号、pct公开案wo94/16716、wo96/39491、wo95/30018;paoletti,“applicationsofpoxvirusvectorstovaccination:anupdate”,pnasusa93:11349-11353,1996年10月;moss,“geneticallyengineeredpoxvirusesforrecombinantgeneexpression,vaccination,andsafety”,pnasusa93:11341-11348,1996年10月;smith等人,美国专利第4,745,051号(重组杆状病毒);richardson,c.d.(编者),methodsinmolecularbiology39,“baculovirusexpressionprotocols”(1995humanapressinc.);smith等人,“productionofhumanbetainterferonininsectcellsinfectedwithabaculovirusexpressionvector”,molecularandcellularbiology,1983年12月,第3卷,第12期,第2156-2165页;pennock等人,“strongandregulatedexpressionofescherichiacolib-galactosidaseininfectcellswithabaculovirusvector”,molecularandcellularbiology1984年3月,第4卷,第3期,第406页;epa0370573;美国申请案第920,197号,1986年10月16日申请;ep专利公开案第265785号;美国专利第4,769,331号(重组疱疹病毒);roizman,“thefunctionofherpessimplexvirusgenes:aprimerforgeneticengineeringofnovelvectors”,pnasusa93:11307-11312,1996年10月;andreansky等人,“theapplicationofgeneticallyengineeredherpessimplexvirusestothetreatmentofexperimentalbraintumors”,pnasusa93:11313-11318,1996年10月;robertson等人,“epstein-barrvirusvectorsforgenedeliverytoblymphocytes”,pnasusa93:11334-11340,1996年10月;frolov等人,“alphavirus-basedexpressionvectors:strategiesandapplications”,pnasusa93:11371-11377,1996年10月;kitson等人,j.virol.65,3068-3075,1991;美国专利第5,591,439号、第5,552,143号;wo98/00166;许可的美国申请案序列号08/675,556及08/675,566,均在1996年7月3日申请(重组腺病毒);grunhaus等人,1992,“adenovirusascloningvectors”,seminarsinvirology(第3卷)第237-52页,1993;ballay等人embojournal,第4卷,第3861-65页,graham,tibtech8,85-87,1990年4月;prevec等人,j.genvirol.70,42434;pctwo91/11525;felgner等人(1994),j.biol.chem.269,2550-2561,science,259:1745-49,1993;及mcclements等人,“immunizationwithdnavaccinesencodingglycoproteindorglycoproteinb,aloneorincombination,inducesprotectiveimmunityinanimalmodelsofherpessimplexvirus-2disease”,pnasusa93:11414-11420,1996年10月;及美国专利第5,591,639号、第5,589,466号及第5,580,859号以及wo90/11092、wo93/19183、wo94/21797、wo95/11307、wo95/20660;tang等人,nature及furth等人,analyticalbiochemistry,尤其与dna表达载体相关。亦参见wo98/33510;ju等人,diabetologia,41:736-739,1998(慢病毒表达系统);sanford等人,美国专利第4,945,050号;fischbach等人(intracel);wo90/01543;robinson等人,seminarsinimmunology第9卷,第271-283页(1997),(dna载体系统);szoka等人,美国专利第4,394,448号(将dna插入至活细胞中的方法);mccormick等人,美国专利第5,677,178号(使用细胞病变病毒);及美国专利第5,928,913号(用于基因递送的载体);以及本文中列举的其他文献。术语“病毒载体”描述一种经遗传修饰的病毒,其通过重组dna技术以一定方式操纵以使其进入宿主细胞,产生特异性生物活性,例如表达由载体携带的转基因。在一特定方面中,转基因为抗原。病毒载体在靶细胞、组织或生物体中可为或可不为复制胜任型。病毒载体的产生可使用本领域所熟知的任何适合的基因工程技术来实现,包括(但不限于)限制性核酸内切酶消化、连接、转化、质粒纯化、dna测序、在细胞培养物中转染的标准技术,如sambrook等人(molecularcloning:alaboratorymanual.coldspringharborlaboratorypress,n.y.(1989))或k.maramorosch及h.koprowski(methodsinvirologyvolumeviii,academicpressinc.london,uk(2014))中所述。病毒载体可并入来自任何已知生物体的基因组的序列。序列可以其天然形式并入或可以任何方式修饰以获得所需活性。举例而言,序列可包含插入、缺失或取代。病毒载体可包括两种或更多种所关注蛋白质的编码区。举例而言,病毒载体可包括第一所关注蛋白质的编码区及第二所关注蛋白质的编码区。第一所关注蛋白质及第二所关注蛋白质可为相同或不同的。在一些实施方式中,病毒载体可包括第三或第四所关注蛋白质的编码区。第三及第四所关注蛋白质可为相同或不同的。由一个病毒载体编码的两种或更多种所关注蛋白质的总长度可变化。举例而言,两种或更多种蛋白质的总长度可为至少约200个氨基酸、至少约250个氨基酸、至少约300个氨基酸、至少约350个氨基酸、至少约400个氨基酸、至少约450个氨基酸、至少约500个氨基酸、至少约550个氨基酸、至少约600个氨基酸、至少约650个氨基酸、至少约700个氨基酸、至少约750个氨基酸、至少约800个氨基酸或更长。优选病毒载体包括疱疹病毒载体,诸如来源于ehv-1或ehv-4或其他水痘病毒属,如prv(伪狂犬病病毒)或bhv-1(牛疱疹病毒1)。根据本发明的特定方面,术语“病毒载体”或“病毒构建体”是指来源于病毒的重组病毒构建体,该病毒是选自疱疹病毒科,诸如ehv-1、ehv-4或其他水痘病毒属,如猪α疱疹病毒1(伪狂犬病病毒,prv)及牛α疱疹病毒1(牛疱疹病毒1,bhv-1);腺病毒科(adv),诸如cadv(犬腺病毒,vanregenmortel等人);细小病毒科,如腺相关病毒;杆状病毒科;反转录病毒科或痘病毒科。(http://www.ictvonline.org/virustaxonomy.aspvirustaxonomy:2015releaseec47,london,uk,2015年7月;emailratification2016(msl#30)。优选病毒载体包括疱疹病毒载体,诸如来源于ehv-1或ehv-4或其他水痘病毒属,如prv(伪狂犬病病毒)或bhv-1(牛α疱疹病毒1)。术语“病毒载体”及“病毒构建体”可互换使用。如本文所用,术语“构建体”是指重组核酸,诸如质粒、bac或已人工产生的重组病毒。术语“质粒”是指独立于细菌宿主细胞内的细菌染色体复制的细胞质dna。在本发明的一特定方面中,术语“质粒”和/或“转移质粒”是指可用于构建例如插入至病毒载体中的表达盒的重组dna技术组件。在另一特定方面中,术语“质粒”可用于限定可用于dna疫苗接种目的的质粒。如本文所用,术语“核酸”及“多核苷酸”可互换且是指任何核酸。如本文所用,术语“核酸”、“核酸序列”、“核苷酸序列”、“多核苷酸”、“多核苷酸序列”、“rna序列”或“dna序列”是指寡核苷酸、核苷酸或多核苷酸及其片段及部分且是指基因组或合成来源的dna或rna,其可为单链或双链且代表有义链或反义链。序列可为非编码序列、编码序列或两者的混合物。本发明的核酸序列可使用本领域技术人员熟知的标准技术制备。术语“核酸”及“多核苷酸”亦特别包括由除五种生物学上存在的碱基(腺嘌呤、鸟嘌呤、胸腺嘧啶、胞嘧啶及尿嘧啶)外的碱基构成的核酸。术语“调控核酸”、“调控组件”及“表达控制组件”可互换使用且是指可影响可操作地连接的编码序列在特定宿主生物体的表达的核酸分子。这些术语使用广泛且涵盖所有促进或调控转录的组件,包括启动子、启动子序列、rna聚合酶及转录因子的碱基相互作用所需的核心组件、上游组件、增强子及反应组件。原核生物中的例示性调控组件包括启动子、操纵子序列及核糖体结合位点。用于真核细胞中的调控组件可包括(但不限于)转录及翻译控制序列,诸如启动子、增强子、剪接信号、聚腺苷酸化信号、终止子、蛋白质降解信号、内部核糖体进入位点(ires)、小核糖核酸病毒科2a序列及其类似物,其提供和/或调控宿主细胞中编码序列的表达和/或经编码多肽的产生。“内部核糖体进入位点”或“ires”描述功能上促进独立于ires5'的基因的翻译开始且允许在动物细胞中由单个转录物翻译两个顺反子(开放阅读框架)的序列。ires提供独立的核糖体进入位点以翻译紧接其下游的开放阅读框架。与可为多顺反子(亦即编码由mrna依序翻译的数个不同多肽)的细菌mrna不同,动物细胞的大多数mrna为单顺反子且编码合成仅一个多肽。关于真核细胞中的多顺反子转录物,翻译将自最5'的翻译起始位点开始,在第一终止密码子处终止,且转录物将自核糖体释放,使得mrna仅翻译第一经编码多肽。在真核细胞中,具有可操作地连接于转录物中的第二或后续开放阅读框架的ires的多顺反子转录物允许下游开放阅读框架的依序翻译,产生由同一转录物编码的两个或更多个多肽。ires可具有不同长度且来自不同来源,例如脑心肌炎病毒(emcv)、小核糖核酸病毒(例如口蹄疫病毒、fmdv或脊髓灰质炎病毒(pv)或c型肝炎病毒(hcv)。本领域已描述且熟知各种ires序列及其在载体构建中的用途。下游编码序列在将不会负面影响下游基因表达的任何距离处可操作地连接于ires的3'端。ires与下游基因起点之间的最优选或可容许的距离可易于通过改变距离且测量随距离而变的表达来确定。术语“2a”或“2a肽”意谓描述为2a及“2a样”的短寡肽序列,其充当能够通过定义为核糖体跳跃的过程介导蛋白质间的共翻译切割的连接子。此类2a及“2a样”序列(来自小核糖核酸病毒科及其他病毒或细胞序列)可用于将多个基因序列串接成单个基因,确保其在同一细胞内共表达(参见luke及ryan,2013)。如本文所用,术语“启动子”或“启动子序列”意谓允许rna聚合酶结合且引导基因转录的核苷酸序列。通常,启动子位于基因的5'非编码区,邻近基因的转录起始位点。用于起始转录的启动子内的序列组件常常通过共同核苷酸序列表征。启动子的实例包括(但不限于)来自细菌、酵母、植物、病毒及动物,诸如哺乳动物(包括马、猪、牛及人类)、鸟类或昆虫的启动子。启动子可为诱导性、阻抑性和/或组成性。诱导性启动子响应于培养条件的一些变化,诸如温度变化,引发在其控制下的dna的转录水平增加(ptashne,2014)。本领域技术人员熟知的启动子的实例为例如sv40大t、hcmv及mcmv立即早期基因1、人类延长因子α启动子、杆状病毒多角体蛋白启动子。如本文所用,在本发明的上下文中,术语启动子尤其是指功能片段,例如截短的4pgg600(seqidno.1)或其互补核苷酸序列,相对于整个长度的序列相同性优选为(至少)72%(或更高)。此外,如本文所用,在本发明的上下文中,术语启动子尤其是指功能片段,例如截短的4pmcp600(seqidno.2)或其互补核苷酸序列,相对于整个长度的序列相同性优选为(至少)78%(或更高)。最优选地,“启动子”是指p430(seqidno.:3)或p455(seqidno.:4)。如本文进一步所用,在本发明的上下文中,术语启动子尤其是指p430(seqidno.:3)或p455(seqidno.:4)或4pgg600(seqidno.1)或4pmcp600(seqidno.2)的功能衍生物,其具有例如少量取代、突变或反转,以使得序列相同性为70%、80%、85%、90%、95%、99%一致或同源。术语“p430”、“gg430”及“430”贯穿本说明书、图式、序列表等同义且可互换地使用。术语“p455”、“mcp455”及“455”贯穿本说明书、图式、序列表等同义且可互换地使用。术语“增强子”表示在顺式位置作用于启动子的活性且因此刺激功能上连接至此启动子的基因或编码序列转录的多核苷酸序列。与启动子不同,增强子的作用与位置及取向无关,且其可因此位于转录单元前面或后面,位于内含子内或甚至位于编码区内。增强子可紧邻转录单元及在距启动子相当大的距离处定位。亦可与启动子具有实体及功能重叠。本领域技术人员应了解来自各种来源的许多增强子(且存储在诸如genbank的数据库中,例如sv40增强子、cmv增强子、多瘤病毒增强子、腺病毒增强子),其以独立组件或在多核苷酸序列中克隆的组件形式得到(例如保藏在atcc或来自商业及个体来源)。许多启动子序列亦含有增强子序列,诸如经常使用的cmv启动子。人类cmv增强子为迄今鉴别的最强的增强子之一。诱导性增强子的一个实例为金属硫蛋白增强子,其可由糖皮质激素或重金属刺激。术语“互补核苷酸序列”描述诸如dna或rna的多核苷酸的两个成对链中的一链。互补链的核苷酸序列反映其成对链的核苷酸序列,使其含有相对于各腺嘌呤的胸腺嘧啶(或对于rna而言,尿嘧啶),相对于各鸟嘌呤的胞嘧啶,反的亦然。例如5'-gcatac-3'的互补核苷酸序列为3'-cgtatg-5'或对于rna而言,3'-cguaug-5'。如本文所用,术语“基因”、“所关注基因”具有相同含义且是指编码所关注产物的任何长度的多核苷酸序列。基因可进一步包含在编码序列之前(5'非编码或非翻译序列)及之前(3'非编码或非翻译序列)的调控序列。所选序列可为全长或截短的融合或加标签基因,且可为cdna、基因组dna或dna片段。大体上了解,编码多肽或rna的基因组dna可包括自成熟信使rna(mrna)剪接的非编码区(亦即内含子)且因此不存在于编码相同多肽或rna的cdna中。其可为天然序列,亦即天然存在的形式,或可突变,或包含来源于不同来源的序列或视需要以其他方式修饰。这些修饰包括密码子优化以使所选宿主细胞中的密码子使用优化或加标签。此外,其可包括移除或添加顺式作用位点,诸如(隐蔽的)剪接供体、受体位点及分支点、聚腺苷酸化信号、tata盒、chi位点、核糖体进入位点、重复序列、二级结构(例如茎环)、转录因子或其他调控因子的结合位点、限制酶位点等,仅给出若干但非限制性的实例。所选序列可编码分泌性、细胞质、细胞核、膜结合或细胞表面多肽。如本文所用,术语“所关注核苷酸序列”为比所关注基因更通用的术语,因为其未必包含基因,但可包含基因的组件或部分或其他基因信息,例如ori(复制起点)。所关注核苷酸序列可为任何dna或rna序列,与其是否包含编码序列无关。“开放阅读框架”或“orf”是指包含翻译起始信号或起始密码子(诸如atg或aug)及终止密码子且可能翻译成多肽序列的一段核酸序列(dna或rna)。术语“转录”描述细胞中mrna的生物合成。如本文所用,术语“表达”是指宿主细胞内核酸序列的转录和/或翻译。根据本发明的特定方面,术语“表达”是指宿主细胞内异源和/或外源核酸序列的转录和/或翻译。宿主细胞中所需产物的表达量可基于细胞中存在的对应rna或mrna的量或由所选序列编码的所需多肽的量来确定。举例而言,自所选序列转录的mrna可通过核酸印迹杂交、核糖核酸酶rna保护、与细胞rna原位杂交或通过rtqpcr(反转录后接定量pcr)来定量。自所选序列表达的蛋白质可通过各种方法,例如通过elisa、通过免疫印迹法、通过放射免疫分析、通过免疫沉淀、通过分析蛋白质的生物活性或通过蛋白质免疫染色后接facs分析来定量。术语“表达盒”或“转录单元”或“表达单元”定义载体、构建体或多核苷酸序列内含有一或多个有待转录的基因的区域,其中表达盒内所含的编码经转录基因的核苷酸序列以及含有调控组件的多核苷酸序列可操作地彼此连接。其自启动子转录且通过至少一个聚腺苷酸化信号终止转录。在一个特定方面中,其自一个单一启动子转录。因此,不同基因至少在转录上相连。可自各转录单元(多顺反子转录单元)转录及表达大于一种蛋白质或产物。各转录单元将包含该单元内所含的任何所选序列的转录及翻译必需的调控组件。且各转录单元可含有相同或不同的调控组件。举例而言,各转录单元可含有同一终止子、ires组件或可用于转录单元内基因的功能性连接的内含子。载体或多核苷酸序列可含有大于一个转录单元。术语“增加的表达”、“增加的效价或生产率”或“改良的表达或生产率”意谓引入至宿主细胞中的异源和/或外源序列、例如编码治疗性蛋白质的基因的表达、合成或分泌与适合的对照物相比(例如由cdna编码的蛋白质对比由含有内含子的基因编码的蛋白质)增加。若根据本发明的细胞根据此处所描述的根据本发明的方法培养,及若此细胞的比生产率或效价具有至少1.2倍、1.5倍、两倍、三倍、四倍或五倍增加,则存在增加的效价或生产率。若根据本发明的细胞根据此处所描述的根据本发明的方法培养,及若此细胞的比生产率或效价具有至少1.2倍或至少1.5倍或至少两倍或至少三倍增加,则亦存在增加的效价或生产率。若根据本发明的细胞根据此处所描述的根据本发明的方法培养,及若此细胞的比生产率或效价具有至少1.2倍至五倍、优选1.5倍至五倍、更优选两倍至五倍、尤其优选三倍至五倍增加,则亦尤其存在增加的效价或生产率。“增加的表达”亦可意谓更多细胞实际上表达所关注基因/序列。举例而言,增加的表达可意味着本发明的新颖的启动子在病毒复制周期期间的活性相对于其他启动子持续较长时间段。增加的表达、效价或生产率可通过使用根据本发明的异源载体来获得。此可与其他方法组合,诸如facs辅助选择含有一或多个荧光蛋白(例如gfp)或细胞表面标记作为额外可选标记的重组宿主细胞。亦可使用的其他获得增加的表达的方法及不同方法的组合是基于例如使用顺式作用组件操控染色质结构(例如lcr、ucoe、ease、隔离子、s/mar、star组件)、使用(人工)转录因子、用天然或合成试剂处理细胞以上调内源或异源和/或外源基因表达、改良mrna或蛋白质的稳定性(半衰期)、改良mrna翻译的起始、通过使用游离型质粒增加基因剂量(基于使用病毒序列作为复制起点,例如sv40、多瘤病毒、腺病毒、ebv或bpv)、使用扩增促进序列或基于dna串联体的活体外扩增系统。测量“增加的表达”的分析为基于lc-ms/ms的蛋白质测量,诸如多反应监测(mrm);基于抗体的检测方法,诸如免疫印迹法、斑点印迹或免疫扩散及流式细胞测量术;及通过血球凝集分析测量生物活性。“启动子活性”通过定量在相应启动子控制下转录的mrna来间接测量。mrna通过rtqpcr相对于内源性标准来定量。术语“病毒效价”为每一体积病毒制剂的感染单位的量度。病毒效价为生物程序的端点且定义为并行进行的特定比例的测试显示效应的稀释度(reed及muench,1938)。具体地,每毫升组织培养物感染剂量五十(tcid50/ml)给出用稀释液并行接种的多个细胞培养物的50%经感染的病毒制剂的稀释度。“转录调控组件”通常包含有待表达的基因序列上游的启动子、转录起始及终止位点以及聚腺苷酸化信号。术语“转录起始位点”是指构建体中对应于并入至初级转录物(亦即mrna前体)中的第一核酸的核酸。转录起始位点可与启动子序列重叠。“终止信号”或“终止子”或“聚腺苷酸化信号”或“聚腺苷酸(polya)”或“转录终止位点”或“转录终止组件”为在真核mrna的3'端的特定位点引起切割及在切割的3'端转录后并入约100-200个腺嘌呤核苷酸序列(聚腺苷酸尾部)且因此致使rna聚合酶终止转录的信号序列。聚腺苷酸化信号包含在切割位点上游约10-30个核苷酸处的序列aataaa及位于下游的序列。已知多种聚腺苷酸化组件,诸如tk聚腺苷酸、sv40后期及早期聚腺苷酸、bgh聚腺苷酸(描述于例如美国专利第5,122,458号中)或仓鼠生长激素聚腺苷酸(wo2010010107)。对于各有待表达的单个多肽,“翻译调控组件”包含翻译起始位点(aug)、终止密码子及聚腺苷酸信号。内部核糖体进入位点(ires)可包括于一些构建体中。为了使表达优化,移除、添加或改变有待表达的核酸序列的5'-和/或3'-非翻译区以消除任何可能附加的不当替代性翻译起始密码子或可能在转录或翻译水平上干扰或减少表达的其他序列可为可取的。共同核糖体结合位点(kozak序列)可紧接着插入起始密码子上游以增强翻译且因此增强表达。此核糖体结合位点周围增加的a/u含量促进更高效的核糖体结合。按照定义,插入于宿主细胞中的每一多核苷酸序列或每一基因及由其编码的相应蛋白质或rna当来自不同(病毒)物种时,相对于宿主细胞称为“外源”、“外源序列”、“外源基因”、“外源编码序列”。因此,本发明的基于ehv-4的启动子鉴于ehv-1病毒载体或cadv病毒载体为外源的。如本文所用,相对于所关注序列或基因,诸如抗原,术语“外源”意谓该所关注序列或基因、具体地该抗原是在其天然物种情形以外表达。因此,来自猪iav的h3抗原为相对于ehv-1载体的外源抗原的一个实例(参见实例3)。所关注的任何非马序列或基因、诸如非马抗原因此为根据本发明的一特定方面的所关注的外源序列或基因或抗原。按照定义,插入于宿主细胞中的每一多核苷酸序列或每一基因及由其编码的相应蛋白质或rna相对于宿主细胞称为“异源”、“异源序列”、“异源基因”、“异源编码序列”、“转基因”或“异源蛋白质”。即使有待引入的序列或有待引入的基因与宿主细胞的内源序列或内源基因一致,其仍适用。举例而言,与ehv-4野生型病毒相比在不同位点或以修饰形式引入至ehv-4病毒载体中的ehv-4启动子序列按照定义为异源序列。如本文所用,相对于所关注序列或基因,诸如抗原,术语“异源”意谓该所关注序列或基因、具体地该抗原是在其天然亚种情形以外表达。因此,所关注的任何非ehv-1特异性序列或基因,诸如抗原,例如来自除ehv-1之外的任何马α疱疹病毒(例如ehv-3、ehv-8)的抗原因此为根据本发明的一特定方面的所关注的异源序列或基因或抗原。术语“非天然存在”意谓在此上下文中并非天然存在的任何所关注序列或基因(诸如抗原),诸如杂交序列或来自不同物种的所关注序列或基因(诸如抗原),或由于人工突变、插入、缺失或其类似物而并非天然产物的所关注序列或基因(诸如抗原)。术语“重组”在本发明的说明书通篇与术语“非天然存在”、“异源”及“外源”可互换使用。因此,“重组”蛋白质为自异源或外源多核苷酸序列表达的蛋白质。如关于病毒所用,术语重组意谓通过人工操纵病毒基因组产生的病毒。包含异源或外源序列、诸如外源抗原编码序列的病毒为重组病毒。术语重组病毒及术语非天然存在的病毒可互换使用。因此,术语“异源载体”意谓包含异源或外源多核苷酸序列的载体。术语“重组载体”意谓包含异源或重组多核苷酸序列的载体。如本文所用,术语“可操作地连接”用于描述调控组件与基因或其编码区之间的连接。通常,基因表达在一或多个调控组件控制下,例如(但不限于)组成性或诱导性启动子、组织特异性调控组件及增强子。基因或编码区称为“可操作地连接于”调控组件或与调控组件“可操作地相关”,意味着该基因或编码区受调控组件控制或影响。举例而言,若启动子影响编码序列的转录或表达,则启动子可操作地连接于编码序列。此外,在本说明书的范畴内,术语“功能性连接”、“功能性地连接”或“可操作地连接”意谓两个或更多个核酸序列或序列组件以允许其按预期方式起作用的方式定位。举例而言,启动子/增强子或终止子若能够在顺式位置控制或调节所连接的基因序列的转录,则功能性地连接于编码基因序列。一般但非必须地,功能性地连接的dna序列为连续的且必要时接合两个多肽编码区,或在分泌信号肽的情况下,为连续的且在阅读框架中。然而,虽然可操作地连接的启动子一般位于编码序列上游或可操作地连接的终止子一般位于编码序列下游,但不必与其邻接。增强子无须邻接,只要其增加编码序列的转录即可。为此,其可位于编码序列上游或下游且甚至在一定距离处。聚腺苷酸化位点若以转录前进经过编码序列至聚腺苷酸化信号的方式位于编码序列的3'端,则可操作地连接于编码序列。连接通过本领域已知的重组方法实现,例如通过在适合的限制性位点或钝端处连接或通过使用融合pcr方法。若不存在适合的限制性位点,则可根据常规实践使用合成寡核苷酸连接子或接头。因此,启动子序列的术语“功能片段”或“功能衍生物”意谓该片段或衍生物仍影响启动子活性。如何评定启动子活性的功能分析为一般本领域技术人员所熟知的(bustin2000,nolan等人2006)。此类功能分析的例示性实施例包括例如启动子动力学实验。培育感染有载体病毒的细胞持续不同时间,该等载体病毒携带表达盒,其中启动子或其片段引导报告子转基因的转录。总rna是由在感染后的不同时间采集的样品制备。在通过dna酶i消化破坏污染性dna之前,使rna反转录。一个复制样品在添加反转录酶(rt)的情况下进行处理,第二复制样品在不添加rt的情况下进行处理,以便证明污染性dna自rna制备物成功移除。将所得cdna纯化且用作常规pcr的模板。仅在添加rt的情况下处理的样品将产生pcr产物。这些cdna可随后与报告子转基因的引物以及病毒载体的必需基因(内标物基因)的引物一起用于qpcr,该内标物基因的转录提供感染及复制效率的内标物。使用内标物基因的qpcr值使报告子的qpcr值在不同构建体与感染后的时间之间标准化。此举允许说明不同启动子及其片段的启动子活性。如本文所用,“序列同源性”是指一种确定两个序列相关性的方法。为确定序列同源性,将两个或更多个序列进行最优选比对,且必要时引入间隙。然而,相比于“序列相同性”,当确定序列同源性时,将保守氨基酸取代视为匹配。换言之,为获得与参考序列具有95%序列同源性的可比多肽或多核苷酸,参考序列中85%、优选90%、91%、92%、93%、94%、甚至更优选95%、96%、97%、98%、99%、99.9%的氨基酸残基或核苷酸必须匹配或包含经另一氨基酸或核苷酸保守取代。或者,可将参考序列中不包括保守取代的总氨基酸残基或核苷酸的至多15%、优选至多10%、9%、8%、7%、6%、甚至更优选至多5%、4%、3%、2%、1%、0.1%的多个氨基酸或核苷酸插入至参考序列中。优选地,同源序列包含至少50、甚至更优选100、甚至更优选250、甚至更优选500个核苷酸的延伸。如本领域已知,“序列相同性”是指两个或更多个多肽序列或两个或更多个多核苷酸序列,亦即参考序列及有待与参考序列比较的给定序列之间的关系。序列相同性通过在序列已最优选比对产生最高程度的序列相似性后比较给定序列与参考序列来确定,如通过此类序列串之间的匹配所确定。在此比对时,序列相同性是在逐个位置基础上来确定,例如若在特定位置处的核苷酸或氨基酸残基一致,则序列在该位置处“一致”。此类位置一致的总数随后除以参考序列中核苷酸或残基的总数,得到序列相同性%。序列相同性可易于通过已知方法计算,包括(但不限于)computationalmolecularbiology,lesk,a.n.,编,oxforduniversitypress,newyork(1988),biocomputing:informaticsandgenomeprojects,smith,d.w.,编,academicpress,newyork(1993);computeranalysisofsequencedata,parti,griffin,a.m.及griffin,h.g.,编,humanapress,newjersey(1994);sequenceanalysisinmolecularbiology,vonheinge,g.,academicpress(1987);sequenceanalysisprimer,gribskov,m.及devereux,j.,编,m.stocktonpress,newyork(1991);及carillo,h.及lipman,d.,siamj.appliedmath.,48:1073(1988)中所述的方法,其教示内容以引用的方式并入本文中。确定序列相同性的优选方法经设计以给出所测试序列之间的最大匹配。确定序列相同性的方法编码在可公开获得的确定给定序列之间的序列相同性的计算机程序中。此类程序的实例包括(但不限于)gcg程序包(devereux,j.,等人,nucleicacidsresearch,12(1):387(1984))、blastp、blastn及fasta(altschul,s.f.等人,j.molec.biol.,215:403-410(1990)。blastx程序可自ncbi及其他来源公开获得(blastmanual,altschul,s.等人,ncvinlmnihbethesda,md20894,altschul,s.f.等人,j.molec.biol.,215:403-410(1990),其教示内容以引用的方式并入本文中)。这些程序使用默认间隙权重最优选比对序列,以产生给定序列与参考序列之间的最高序列相同性水平。通过核苷酸序列与参考核苷酸序列具有至少例如85%、优选90%、91%、92%、93%、94%、甚至更优选95%、96%、97%、98%、99%、99.9%“序列相同性”的多核苷酸作为说明,意欲除给定多核苷酸序列可包括参考核苷酸序列的每100个核苷酸至多15、优选至多10、甚至更优选至多5个点突变之外,给定多核苷酸的核苷酸序列与参考序列一致。换言之,在核苷酸序列相对于参考核苷酸序列具有至少85%、优选90%、91%、92%、93%、94%、甚至更优选95%、96%、97%、98%、99%、99.9%相同性的多核苷酸中,参考序列中至多15%、优选10%、9%、8%、7%、6%、甚至更优选5%、4%、3%、2%、1%、0.1%的核苷酸可缺失或经另一个核苷酸取代,或参考序列中总核苷酸的至多15%、优选10%、9%、8%、7%、6%、甚至更优选5%、4%、3%、2%、1%、0.1%的多个核苷酸可插入至参考序列中。参考序列的这些突变可发生于参考核苷酸序列的5'或3'末端位置或那些末端位置之间的任何位置,这些位置个别地散置于参考序列的核苷酸中或参考序列内的一或多个邻接基团中。类似地,通过给定氨基酸序列与参考氨基酸序列具有至少例如85%、优选90%、91%、92%、93%、94%、甚至更优选95%、96%、97%、98%、99%序列相同性的多肽,意欲除给定多肽序列可包括参考氨基酸序列的每100个氨基酸至多15、优选至多10、9、8、7、6、甚至更优选至多5、4、3、2、1个氨基酸变化之外,多肽的给定氨基酸序列与参考序列一致。换言之,为获得与参考氨基酸序列具有至少85%、优选90%、91%、92%、93%、94%、甚至更优选95%、96%、97%、98%、99%序列相同性的给定多肽序列,参考序列中至多15%、优选至多10%、9%、8%、7%、甚至更优选至多5%、4%、3%、2%、1%的氨基酸残基可缺失或经另一个氨基酸取代,或参考序列中氨基酸残基总数的至多15%、优选至多10%、9%、8%、7%、甚至更优选至多5%、4%、3%、2%、1%的多个氨基酸可插入至参考序列中。参考序列的这些改变可发生于参考氨基酸序列的氨基或羧基末端位置或那些末端位置之间的任何位置,这些位置个别地散置于参考序列的残基中或参考序列内的一或多个邻接基团中。不一致残基位置的差异优选为保守氨基酸取代。然而,当确定序列相同性时,保守取代不包括为匹配。术语“序列相同性”或“相同性百分比”在本文中可互换使用。出于本发明的目的,此处规定为了确定两个氨基酸序列或两个核酸序列的相同性百分比,出于最优选比较目的比对序列(例如可在第一氨基酸或核酸序列中引入间隙以便与第二氨基酸或核酸序列最优选比对)。接着比较相应氨基酸位置或核苷酸位置处的氨基酸残基或核苷酸残基。当第一序列中的位置与第二序列中的相应位置由相同氨基酸残基或核苷酸残基占据时,则分子在该位置处一致。两个序列之间的相同性百分比为该等序列共有的一致位置数的函数(亦即,相同性%=一致位置数/位置总数(亦即重叠位置)×100)。优选地,两个序列的长度相同。可在所比较的两个序列的整个长度上或在两个序列的片段上进行序列比较。通常,将在所比较的两个序列的全长上进行比较。然而,序列相同性可在例如二十个、五十个、一百个或更多个邻接氨基酸残基的区域上进行。技术人员应了解数个不同计算机程序可用于确定两个序列之间的同源性。举例而言,可使用数学算法进行序列比较及测定两个序列之间的相同性百分比。在一优选实施例中,使用已并入accelrysgcg软件包中的gap程序(可在http://www.accelrys.com/products/gcg/获得)中的needleman及wunsch(j.mol.biol.(48):444-453(1970))算法,使用blosum62矩阵或pam250矩阵及16、14、12、10、8、6或4的间隙权重及1、2、3、4、5或6的长度权重,确定两个氨基酸或核酸序列之间的相同性百分比。技术人员应了解,所有这些不同参数将产生略微不同的结果,但两个序列的总体相同性百分比在使用不同算法时不显著改变。本发明的蛋白质序列或核酸序列可进一步用作“查询序列”,以针对公共数据库进行搜寻,例如以鉴别其他家族成员或相关序列。此类搜寻可使用altschul等人(1990)j.mol.biol.215:403-10的blastn及blastp程序(2.0版)进行。blast蛋白质搜寻可用blastp程序、得分=50、字长=3进行,从而获得与本发明的蛋白质分子同源的氨基酸序列。为实现间隙比对以达成比较目的,可如altschul等人(1997)nucleicacidsres.25(17):3389-3402中所述使用间隙blast。当利用blast及间隙blast程序时,可使用相应程序(例如blastp及blastn)的预设参数。参见美国国家生物技术信息中心(thenationalcenterforbiotechnologyinformation)在http://www.ncbi.nlm.nih.gov/的主页。ehv-1及ehv-4/重组载体技术定义术语“马”意谓马科的或属于马科,其包括马、驴及斑马,优选为马。另外,术语“马”亦涵盖马科成员的杂种(例如骡等)。“疱疹病毒”或“疱疹病毒载体”是指疱疹病毒目疱疹病毒科中的物种。术语“马疱疹病毒载体”或“马疱疹病毒”或“ehv”意谓侵袭马的疱疹病毒科成员。迄今已鉴别马疱疹病毒的八种不同物种,五种属于α疱疹病毒亚科(ehv-1、ehv-3、ehv-4、ehv-8及ehv-9)且三种属于γ疱疹病毒科。(http://www.ictvonline.org/virustaxonomy.aspvirustaxonomy:2015releaseec47,london,uk,2015年7月;emailratification2016(msl#30)术语“ehv-1”意谓疱疹病毒科α疱疹病毒属中水痘病毒亚属的成员马α疱疹病毒1。ehv-1的非限制性参考序列应为例如野生型ehv-1病毒株ab4(genbank登陆编号ay665713.1)或rach(hübert1996)。术语ehv-4意谓疱疹病毒科α疱疹病毒属中水痘病毒亚属的成员马α疱疹病毒4。术语“cadv”或“cav”或“cav2”或“cav-2”是指腺病毒科哺乳动物腺病毒属的成员2型犬腺病毒。以前,术语犬腺病毒1(cav-1或cav1)及犬腺病毒2(cav-2或cav2)用于指定哺乳动物腺病毒的2个不同物种。然而,根据较新分类(http://www.ictvonline.org/virustaxonomy.aspvirustaxonomy:2015releaseec47,london,uk,2015年7月;emailratification2016(msl#30),术语犬腺病毒(cadv)现在涵盖物种cav-2及cav-1。术语“插入至orf70中”意谓dna片段在编码马α疱疹病毒1开放阅读框架70的位置处插入至基因组dna中。在本发明的一特定方面中,所提及的插入致使orf70的801个5'碱基对缺失,保留3'端的其余423bp不变,但消除orf70基因产物糖蛋白g的表达。已展示包括ehv-1的数个α疱疹病毒的糖蛋白g自经感染细胞分泌且通过结合促炎性细胞介素充当免疫调节蛋白质。相比于具有完整糖蛋白g表达的野生型ehv-1,在病毒载体中消除糖蛋白g的表达应增加病毒感染的免疫原性。术语“插入至orf1/3中”意谓dna片段在一位置处插入于病毒基因组中,该位置在疫苗株ehv-1rach的减毒程序期间,因传代中的意外缺失而失去包含90%orf1及整个orf2的1283bp片段。选择此插入位点是因为预计来自此位置的转基因的表达将干扰病毒复制的可能性极低。疫苗定义“免疫原性或免疫组合物”是指包含至少一个抗原或其免疫原性部分的物质的组合物,其在宿主体内引发对该组合物的细胞免疫反应或抗体介导的免疫反应的免疫反应。本文所用的术语“抗原”为本领域充分理解的且包括具有免疫原性的物质(亦即免疫原)以及诱发免疫无反应性或失能(亦即缺失身体防御机制对外来物质的反应)的物质。如本文所用,术语“抗原”欲意谓含有或包含表位的全长蛋白质以及其肽片段。术语“产食性动物”意谓给人类食用消费的动物,诸如猪、牛、家禽、鱼类及其类似物,产食性动物优选指猪及牛,最优选为猪。术语“产食性动物”不包括马科动物,诸如马。如本文所用,“免疫原性组合物”是指引发如本文所述的免疫反应的多肽或蛋白质,诸如病毒表面蛋白质。术语“免疫原性片段”或“免疫原性部分”是指包括一或多个表位且因此引发本文所述的免疫反应的蛋白质或多肽的片段或截短和/或取代形式。一般而言,此类截短和/或取代形式或片段包含来自全长蛋白质的至少六个邻接氨基酸。此类片段可使用本领域熟知的多种表位作图技术鉴别。参见例如epitopemappingprotocolsinmethodsinmolecularbiology,第66卷(glenne.morris编,1996)humanapress,totowa,newjersey。举例而言,线性表位可如下测定:通过在固体支持物上并行合成大量肽,该等肽对应于蛋白质分子的部分,且在肽仍附接于支持物时,使肽与抗体反应。此类技术为已知的且描述于本领域,参见例如美国专利第4,708,871号;geysen等人(1984)proc.natl.acad.sci.usa81:3998-4002;及geysen等人(1986)molec.immunol.23:709-715。类地似,可轻易地通过诸如利用例如x射线结晶学及二维核磁共振判定氨基酸的空间构象来鉴别构象表位。参见epitopemappingprotocols,见上文。合成抗原亦包括在该定义内,例如多表位、侧接表位及其他重组或合成衍生抗原。参见例如bergmann等人(1993)eur.j.immunol.23:2777-2781;bergmann等人(1996),j.immunol.157:3242-3249;suhrbier,a.(1997),immunol.andcellbiol.75:402-408;及gardner等人,(1998)12thworldaidsconference,geneva,switzerland,1998年6月28日-7月3日。(其教示及内容全部以引用的方式并入本文中。)如本文所用,术语“疫苗”是指一种医药组合物,该组合物包含至少一种可诱发动物体内免疫反应的免疫活性组分,及一或多种可能但不必然会增强活性组分的免疫活性的额外组分。疫苗可另外包含医药组合物典型的其他组分。作为区别,疫苗的免疫活性组分可包含完整病毒颗粒,他们为原始形式或为所谓改良式活毒疫苗(modifiedlivevaccine;mlv)的减毒颗粒,或所谓死疫苗(killedvaccine;kv)中通过适当方法的失活颗粒。在另一形式中,痘苗的免疫活性组分可包含生物体的适当要素(亚单位疫苗),其中这些要素通过以下产生:破坏整个颗粒或含有此类颗粒的生长培养基且可选地随后进行纯化步骤,产生所需结构;或合成过程,包括通过使用基于例如细菌、昆虫、哺乳动物或其他物种的适合系统进行适当操作加上可选地存在的后续分离及纯化程序;或使用适合医药组合物直接并入遗传物质,在需要疫苗的动物中诱发合成过程(多核苷酸疫苗接种)。疫苗可包含一种或同时超过一种上述要素。如在本发明的特定方面内所用,“疫苗”是指活疫苗或活病毒,亦称为重组疫苗。在本发明的另一特定方面中,“疫苗”是指失活病毒或死病毒,包括病毒样颗粒(vlp)。因此,疫苗可为亚单位疫苗或死疫苗(kv)或失活疫苗。术语“感染复数(m.o.i.)”描述每一细胞使用病毒制剂有多少感染单位(例如tcid50)可感染所培养细胞。举例而言,0.01的m.o.i.意谓培养容器中每100个细胞接种一个感染单位。术语“dna疫苗接种”或“多核苷酸疫苗接种”意谓使用适合的医药组合物直接接种遗传物质。本领域已知失活的各种物理及化学方法。术语“失活”是指先前毒力或非毒力病毒或细菌已经辐照(紫外(uv)、x射线、电子束或γ辐射)、加热或化学处理以使此类病毒或细菌失活或杀死此类病毒或细菌,同时保留其免疫原性。适合的失活剂包括β-丙内酯、二或β-或乙酰基-乙撑亚胺、戊二醛、臭氧及福尔马林(formalin)(甲醛)。对于通过福尔马林或甲醛失活而言,甲醛通常与水及甲醇混合,产生福尔马林。添加甲醇防止在失活过程期间产生降解或交叉反应。一个实施方式使用约0.1%至1%的37%甲醛溶液以使病毒或细菌失活。关键是调节福尔马林的量以确保使物质失活,但不会达到因高剂量引起副作用的程度。更具体地,术语“失活”在病毒的情形下意谓病毒不能活体内或活体外复制,且对应地,术语“失活”在细菌的情形下意谓细菌不能活体内或活体外再生。举例而言,术语“失活”可指病毒已活体外繁殖且随后已使用化学或物理方式失活,从而使其不再能够复制。在另一实例中,术语“失活”可指细菌已繁殖且随后使用化学或物理方式失活,产生细菌、细菌的片段或组分的悬浮液,诸如产生可用作疫苗组分的菌苗。如本文所用,术语“失活”、“死”或“kv”可互换使用。术语“活疫苗”是指包含活生物体或复制胜任型病毒或病毒载体的疫苗。“医药组合物”基本上由一或多种能够改良其施用的生物体或存活在该生物体中或该生物体上的生物体的生理、例如免疫功能的成分组成。该术语包括(但不限于)抗生素或抗寄生虫剂,以及常用以达成某些其他目标的其他成分,该等目标诸如(但不限于)加工性状、无菌性、稳定性、经由经肠或非经肠途径(诸如经口、鼻内、静脉内、肌肉内、皮下、皮内或其他适合途径)施用组合物的可行性、施用后耐受性或控制释放特性。仅出于示范目的而给出的此类医药组合物的一个非限制性实例可如下制备:将经感染细胞培养物的细胞培养上清液与稳定剂(例如亚精胺和/或牛血清白蛋白(bsa))混合且随后通过其他方法将混合物冻干或脱水。在疫苗接种前,混合物接着在水溶液(例如生理食盐水、磷酸盐缓冲生理食盐水(pbs))或非水溶液(例如油乳液、铝类佐剂)中再水合(rehydrate)。如本文所用,“医药学或兽医学上可接受的载剂”包括任何及所有溶剂、分散介质、包衣、佐剂、稳定剂、稀释剂、防腐剂、抗细菌剂及抗真菌剂、等张剂、吸附延迟剂及其类似物。在一些优选实施方式及尤其包括冻干免疫原性组合物的实施方式中,用于本发明的稳定剂包括用于冻干或冷冻干燥的稳定剂。在一些实施方式中,本发明的免疫原性组合物含有佐剂。如本文所用,“佐剂”可包括氢氧化铝及磷酸铝、皂苷,例如quila、qs-21(cambridgebiotechinc.,cambridgema)、gpi-0100(galenicapharmaceuticals,inc.,birmingham,al)、油包水乳液、水包油乳液、水包油包水乳液。乳液可尤其基于轻液态石蜡油(欧洲药典类型);类异戊二烯油,诸如角鲨烷或角鲨烯;由烯烃、尤其异丁烯或癸烯寡聚产生的油;含有直链烷基的酸或醇的酯,更尤其植物油、油酸乙酯、丙二醇二-(辛酸酯/癸酸酯)、甘油基三-(辛酸酯/癸酸酯)或丙二醇二油酸酯;分支链脂肪酸或醇的酯,尤其异硬脂酸酯。油与乳化剂组合使用以形成乳液。乳化剂优选为非离子界面活性剂,尤其脱水山梨糖醇、二缩甘露醇(例如无水甘露糖醇油酸酯)、乙二醇、聚丙三醇、丙二醇及油酸、异硬脂酸、蓖麻油酸或羟基硬脂酸的酯,其可选地经乙氧基化;及聚氧化丙烯-聚氧化乙烯共聚物嵌段、尤其普洛尼克(pluronic)产品、尤其l121。参见hunter等人,thetheoryandpracticalapplicationofadjuvants(ed.stewart-tull,d.e.s.),johnwileyandsons,ny,第51-94页(1995)及todd等人,vaccine15:564-570(1997)。例示性佐剂为描述于“vaccinedesign,thesubunitandadjuvantapproach”,m.powell及m.newman编,plenumpress,1995的第147页上的spt乳液及描述于此同一本书第183页上的乳液mf59。佐剂的另一实例为选自丙烯酸或甲基丙烯酸的聚合物及顺丁烯二酸酐与烯基衍生物的共聚物的化合物。有利的佐剂化合物为尤其与糖或多元醇的聚烯基醚交联的丙烯酸或甲基丙烯酸的聚合物。这些化合物由术语卡波姆(carbomer)已知(pharmeuropa,第8卷,第2期,1996年6月)。本领域技术人员亦可参考美国专利第2,909,462号,其描述与具有至少3个羟基、优选不超过8个羟基的聚羟基化合物交联的此类丙烯酸聚合物,至少三个羟基的氢原子经具有至少2个碳原子的不饱和脂族基团置换。优选基团为含有2至4个碳原子的那些基团,例如乙烯基、烯丙基及其他烯系不饱和基团。不饱和基团本身可含有其他取代基,诸如甲基。以为名销售的产品(bfgoodrich,ohio,usa)尤其适合。其与烯丙基蔗糖或烯丙基异戊四醇交联。尤其可提及carbopol974p、934p及971p。最优选为使用971p。在顺丁烯二酸酐与烯基衍生物的共聚物中的为共聚物ema(monsanto),其为顺丁烯二酸酐与乙烯的共聚物。这些聚合物溶解于水中产生酸溶液,其将优选中和至生理ph值,得到将并入免疫原性、免疫或疫苗组合物本身的佐剂溶液。其他适合的佐剂尤其包括(但不限于)ribi佐剂系统(ribiinc.)、嵌段共聚物(cytrx,atlantaga)、saf-m(chiron,emeryvilleca)、单磷酰基脂质a、阿夫立定(avridine)脂质-胺佐剂、来自大肠杆菌的热不稳定肠毒素(重组或以其他方式)、霍乱毒素、ims1314或胞壁酰二肽、或天然存在或重组的细胞介素或其类似物或内源性细胞介素释放的刺激剂。预期佐剂可以每剂约100μg至约10mg的量、优选以每剂约100μg至约10mg的量、更优选以每剂约500μg至约5mg的量、甚至更优选以每剂约750μg至约2.5mg的量且最优选以每剂约1mg的量添加。或者,佐剂可以最终产物的体积计在约0.01%至50%的浓度下、优选在约2%至30%的浓度下、更优选在约5%至25%的浓度下、再更优选在约7%至22%的浓度下且最优选在10%至20%的浓度下。“稀释剂”可包括水、生理食盐水、右旋糖、乙醇、甘油及其类似物。等张剂可尤其包括氯化钠、右旋糖、甘露糖醇、山梨糖醇及乳糖。稳定剂尤其包括白蛋白及乙二胺四乙酸(edta)的碱金属盐。“经分离”意谓自其天然状态“经人工”改变,亦即若其存在于自然界中,则其已变化或自其原始环境移出或两者。举例而言,天然存在于活生物体中的多核苷酸或多肽未“经分离”,但与其天然状态的共存物质分离的相同多核苷酸或多肽“经分离”,如该术语在本文中所用。“减毒”意谓降低病原体毒力。在本发明中,“减毒”与“无毒”同义。在本发明中,减毒病毒为毒力已降低以使其不引起感染临床征象,但能够在目标动物体内诱发免疫反应的病毒,但亦可意谓与感染非减毒病毒或病原体且未接受减毒病毒的动物的“对照组”相比,临床征象在感染减毒病毒、尤其所主张的ehv-1rach病毒载体的动物中的发生率或严重程度降低。在此上下文中,术语“降低”意谓与如上文所定义的对照组相比,减少至少10%、优选25%、甚至更优选50%、再更优选60%、甚至更优选70%、再更优选80%、甚至更优选90%且最优选100%。因此,减毒、无毒病原体,诸如所主张的减毒病毒载体、尤其所主张的ehv-1(优选地rach)病毒载体适于产生改良式活毒疫苗(mlv)或改良式活免疫原性组合物。在本文中,“有效剂量”意谓(但不限于)抗原在抗原施用的动物体内引发或能够引发免疫反应,使得临床症状减少的量。如本文所用,在组合物的情形下,术语“有效量”意谓免疫原性组合物在动物体内能够诱发免疫反应,降低感染或发生疾病的发生率或减轻严重程度的量。具体地,有效量是指每剂的菌落形成单位(cfu)。或者,在疗法的情况下,术语“有效量”是指疗法足以降低或减轻疾病或病症或其一或多种症状的严重程度或持续时间,阻止疾病或病症进展,引起疾病或病症消退,防止与疾病或病症相关的一或多种症状复发、显现、发作或进展,或增强或改善另一疗法或治疗剂的预防或治疗的量。“免疫反应”意谓(但不限于)出现针对所关注的(免疫原性)组合物或疫苗的细胞免疫反应和/或抗体介导的免疫反应。通常,免疫反应包括(但不限于)一或多个以下效应:产生或活化特异性针对所关注组合物或疫苗中所包括的抗原的抗体、b细胞、辅助t细胞、抑制t细胞和/或细胞毒性t细胞。优选地,宿主将呈现治疗性或保护性免疫(记忆)反应,从而对新感染的抗性将增强和/或疾病的临床严重程度将降低。此类保护作用可通过以下证实:症状数目减少、症状严重程度降低、或不存在与病原体感染相关的一或多种症状、病毒血症发作延迟、病毒续存降低、整体病毒负荷降低和/或病毒排出减少。“对抗疾病的保护”、“保护性免疫”、“功能性免疫”、“减少临床症状”、“诱导/产生中和抗体和/或血清转换”及类似片语意谓通过施用一或多种本发明的治疗性组合物或其组合产生的对抗疾病或病况的部分或完全反应,使得有害效应与在已暴露于疾病或感染的未经免疫个体中所预期相比较少。亦即,在经疫苗接种的个体中感染的有害效应的严重程度减轻。在经疫苗接种的个体中感染可减少、减缓或可能完全预防。在本文中,在意谓完全预防感染的情况下,对其进行特别陈述。若未陈述完全预防,则该术语包括部分预防。在本文中,“降低临床征象的发生率和/或严重程度”或“减少临床症状”意谓(但不限于)与野生型感染相比,减少一组中感染个体的数目、减少或消除展现感染临床征象的个体的数目、或降低一或多个个体中存在的任何临床征象的严重程度。举例而言,其应是指病原体负荷、病原体排出的任何减少、病原体传播减少或有疟疾症状的任何临床征象减少。优选地,与未接受组合物且感染的个体相比,接受本发明的治疗性组合物的一或多个个体中这些临床征象减少至少10%。更优选地,接受本发明的组合物的个体中临床征象减少至少20%,优选至少30%,更优选至少40%,且甚至更优选至少50%。术语“增加的保护”在本文中意谓(但不限于)相对于未经疫苗接种的个体对照组,在经疫苗接种的个体组中与受感染物感染相关的一或多种临床症状统计学上显著减少。术语“临床症状统计学上显著减少”意谓(但不限于)在经以感染物攻击后,经疫苗接种的个体组中至少一种临床症状的发生频率比未经疫苗接种的对照组中低至少10%、优选20%、更优选30%、甚至更优选50%及甚至更优选70%。“持久保护”应是指持续至少3周、但更优选至少3个月、再更优选至少6个月的“改良功效”。就家畜而言,最优选持久保护应持续直至动物以肉出售的平均年龄。术语由病毒诱发的“病毒血症减少”意谓(但不限于)减少进入动物血流的病毒,其中与未接受组合物且可能感染的动物相比,接受本发明的组合物的动物的血清中病毒血症程度(亦即每毫升血清的病毒dna或rna拷贝数或每分升血清的空斑形成菌落数目)减少至少50%。更优选地,接受本发明的组合物的动物中病毒血症程度减少至少90%、优选至少99.9%、更优选至少99.99%且甚至更优选至少99.999%。如本文所用,术语“病毒血症”具体理解为病毒颗粒在动物(尤其哺乳动物、鸟或昆虫)血流中再生和/或循环的病况。“安全性”是指在疫苗接种后经疫苗接种的动物中无以下不良后果,包括(但不限于):基于病毒的疫苗可能恢复至毒力、临床上显著的副作用,诸如持续全身疾病或疫苗施用部位不可接受的炎症。如本文所用,术语“疫苗接种”或其变化形式意谓(但不限于)包括施用本发明的免疫原性组合物的过程,当向动物施用本发明的免疫原性组合物时直接或间接引发或能够引发该动物的免疫反应。在本发明的上下文中,“死亡率”是指由感染引起的死亡,且包括感染严重至使动物安乐死以防痛苦且人为终止其生命的情况。制剂施用组合物的个体优选为动物,包括(但不限于)牛、马、绵羊、猪、家禽(例如鸡)、山羊、猫、犬、仓鼠、小鼠及大鼠,哺乳动物最优选为猪。本发明的制剂包含有效免疫量的一或多种免疫原性组合物及生理学上可接受的媒剂。疫苗包含有效免疫量的一或多种免疫原性组合物及生理学上可接受的媒剂(vehicle)。制剂应适合施用模式。必要时,免疫原性组合物亦可含有少量湿润剂或乳化剂,或ph缓冲剂。免疫原性组合物可为液体溶液、悬浮液、乳液、片剂、丸剂、胶囊、持续释放制剂或粉末。经口制剂可包括标准载剂(carrier),诸如医药级的甘露醇、乳糖、淀粉、硬脂酸镁、糖精钠、纤维素、碳酸镁等。治疗方法优选施用途径包括(但不限于)鼻内、经口、皮内及肌肉内。最优选以单次剂量在饮用水中施用为合乎需要的。本领域技术人员应认识到,本发明的组合物亦可以一次、两次或更多次剂量以及通过其他施用途径施用。举例而言,此类其他途径包括皮下、皮内、腹膜内、皮内且视所需持续时间及治疗有效性而定,根据本发明的组合物可施用一次或亦可间歇地施用数次、例如在每日基础上施用数天、数周或数月及以不同剂量施用,诸如约103至108tcid50(参见上文病毒效价)。在本发明的一特定方面中,剂量为约103至108tcid50,尤其对于活病毒/活疫苗而言。必要时,组合物可存在于包装或分配器装置中,该包装或分配器装置可含有一或多个含有活性成分的单位剂型。包装可例如包含金属或塑料箔,诸如泡壳包装。包装或分配器装置可附有优选关于施用哺乳动物、尤其猪的施用说明书。与此类容器相关联的可为由管理医药或生物产品的制造、使用或销售的政府机构所规定形式的注意事项,该注意事项反映由人类施用的制造、使用或销售机构的批准。序列概述:以下序列特此详述及揭示于本发明中:启动子:seqidno:1ehv-4600bp脱氧核醣核酸序列4pgg600seqidno:2ehv-4600bp脱氧核醣核酸序列4pmcp600seqidno:3ehv-4430bp脱氧核醣核酸序列pg430seqidno:4ehv-4449bp脱氧核醣核酸序列p455seqidno:5特异性针对orf72的第1130号引物seqidno:6特异性针对orf72的第1131号引物seqidno:7特异性针对mcherry的第1079号引物seqidno:8特异性针对mcherry的第1080号引物插入位点:seqidno:9orf70插入区的人工序列核酸pcr引物1017seqidno:10orf70插入区的人工序列核酸pcr引物1018seqidno:11orf1/3插入区的人工序列核酸pcr引物1007seqidno:12orf1/3插入区的人工序列核酸pcr引物1008seqidno:13左(up70)侧接区(417bp)seqidno:14右(up71)侧接区(431bp)seqidno:15野生型ehv-1病毒株ab4(genbank登陆编号ay665713.1)中位于核苷酸127264-127680处的左侧接区(orf70上游)seqidno:16野生型ehv-1病毒株ab4(genbank登陆编号ay665713.1)中位于核苷酸128484-128913处的右侧接区(orf71上游)seqidno:17red系统中截短的侧接区:左(up70)侧接区(283bp)=与417bp“经典”侧接区的3'283bp一致seqidno:18red系统中截短的侧接区:右(up71)侧接区(144bp)=与431bp“经典”侧接区的5'144bp一致seqidno:19野生型ab4(genbank登陆编号ay665713.1)基因组序列中的缺失部分,nt127681-128482seqidno:20rach基因组序列中的缺失部分(因为尚未知晓完整基因组序列,所以无法获得nt编号)质粒/载体序列:seqidno:21转移质粒pu-mc70-bgh的核苷酸序列seqidno.:22转移载体pu70-p455-71k71的核苷酸序列seqidno.:23转移质粒pu70-p455-h3-71k71的核苷酸序列seqidno.:24转移载体pu-1-3-p430-bghkbgh的核苷酸序列seqidno.:25转移质粒pu1-3-p430-h1av-bghkbgh的核苷酸序列血球凝集素序列seqidno:26血球凝集素[a型流感病毒(a/猪/意大利/116114/2010(h1n2))]genbank:adr01746.1h1pdmseqidno:27血球凝集素[a型流感病毒(a/猪/意大利/7680/2001(h3n2))]genbank:abs50302.2h3:seqidno:28血球凝集素[a型流感病毒(a/猪/gent/132/2005(h1n1))]genbank:afr76623.1h1av:seqidno:29血球凝集素[a型流感病毒(a/猪/意大利/4675/2003(h1n2))]genbank:adk98476.1*h1hu*请注意,氨基酸531(x,终止密码子由本发明人改为i):sbv构建体序列seqidno:30gs连接子序列seqidno:31包括用于亚克隆的限制性位点的合成dna序列seqidno:32用于red重组产生prach-se-70-455-sbvgc的dna片段seqidno:33up70f引物seqidno:34up71r引物seqidno:35seq455-f1引物seqidno:36sbvgcf1引物seqidno:37sbvgcr1引物附图说明以下图式形成本说明书的一部分且包括在此以进一步展示本发明的某些方面。参照这些图式中的一或多者,结合本文中呈现的特定实施例的详细描述,可更好地理解本发明。图1.orf70插入位点的示意图ul=特有长区段us=特有短区段ir=内部反向重复序列tr=末端反向重复序列gg=糖蛋白ggpii=糖蛋白iiorf=开放阅读框架bp=碱基对。图2.比较野生型(wt)ehv-1病毒株ab4及减毒疫苗株ehv-1rach的orf1/3区的示意性图解。图3.启动子动力学实验的qpcr结果。3.a中的图形展示编码必需糖蛋白d的orf72的转录动力学。这些数据用于使mcherry的转录动力学数据(3.b中的图形)标准化。图4.两个独立启动子动力学实验的qpcr结果:描绘转录活性与值的正相关性。将在感染后不同时间的mcherryqpcr结果的标准化ct值自t=0时的相应平均ct值减去。展示两个在两种不同细胞株中的实验。图5.用于将表达盒p455-h3-71插入至ehv-1rach的orf70中的转移质粒的质粒图谱。3'端orf69侧接上游插入位点的病毒基因组dna序列3'端orf70侧接下游插入位点的病毒基因组dna序列p455驱动转基因表达的启动子h3转基因(iav血球凝集素)71pa聚腺苷酸化序列i-sce1i-sce1的切割位点启动子aph驱动卡那霉素抗性基因表达的原核启动子kana卡那霉素抗性orfori质粒载体的复制起点apr安比西林抗性基因ecori、sali、noti、hindiii、kpni、bamhi、xbai指示限制性核酸内切酶切割位点图6.放大orf70插入区的rehv-1rach-se-70-p455-h3基因组的示意性图解。orf69:orf70中插入位点上游的第69号开放阅读框架;p455:本文所述的新颖的启动子,参见例如实施例1;h3:转基因流感病毒血球凝集素;71pa:新颖的聚腺苷酸化序列;δorf70:含有orf71的启动子的orf70的其余部分,其编码结构性病毒糖蛋白ii(gpii)。图7.间接免疫荧光分析:经rehv-1rach-se-70-p455-h3感染的vero-细胞的间接免疫荧光分析24hp.i.细胞用乙醇固定且风干。使用针对h3的商业单克隆抗体作为初级抗体及fitc结合的兔抗小鼠igg作为二级抗体,通过荧光显微术展示经重组ehv-1rachse-70-p455-h3感染的细胞中的h3。图8.免疫印迹法:经不同传代的rehv-1rach-se-70-p455-h3或对照物rehv-1rach-se感染或模拟感染的细胞的免疫印迹法。左侧的印迹用针对ehv-1的gpii的单克隆抗体ai2g7孵育。右侧的复制印迹用针对a型流感血球凝集素h3的商业兔高免疫血清(pa5-34930)孵育。条带的附注:1:经rehv-1rach-se-70-p455-h3p5感染的细胞2:经rehv-1rach-se-70-p455-h3p10感染的细胞3:经rehv-1rach-se-70-p455-h3p15感染的细胞4:经rehv-1rach-se-70-p455-h3p20感染的细胞5:经rehv-1rach-mc70感染的细胞6:未经感染的细胞。图9a及9b.病毒效价:图形展示经疫苗接种或未经疫苗接种的猪的肺样品在攻击后的病毒负荷inact=市售失活疫苗ehv=rehv-1rach-se-70-p455-h3。图10.用于将表达盒p430-h1av-bgh插入至ehv-1rach的orf1/3中的转移质粒的质粒图谱。侧接序列a侧接上游插入位点的病毒基因组dna序列侧接序列b侧接下游插入位点的病毒基因组dna序列p430驱动转基因表达的启动子h1av转基因(iav血球凝集素)bghpa聚腺苷酸化序列i-sce1i-sce1的切割位点启动子aph驱动卡那霉素抗性基因表达的原核启动子kana卡那霉素抗性orfori质粒载体的复制起点apr安比西林抗性基因ecori、sali、noti、hindiii、kpni、bamhi指示限制性核酸内切酶切割位点图11.放大orf1/3插入区的rehv-1rach-se-1/3-p430-h1av基因组的示意性图解。δorf1:插入位点上游的开放阅读框架1的其余部分;p430:本文所述的新颖的启动子,参见例如实例1;h1av:转基因流感病毒血球凝集素;bghpa:牛生长激素聚腺苷酸化序列;orf3:插入位点下游的开放阅读框架3。图12.经rehv-1rach-se-1/3-p430-h1av感染的细胞的免疫印迹法及免疫荧光展示转基因的表达。h1av=rehv-1rach-se1/3-p430-h1avse=rehv-rach-se(对照物)模拟物=未感染细胞(对照物)图13.放大两个插入区的rehv-1rach-se-1/3-p430-h1av-70-p455-h3(rehv-1-rach-seb)基因组的示意性图解。δorf1:插入位点上游的开放阅读框架1的其余部分;p430:新颖的启动子;h1av:转基因流感病毒血球凝集素;bghpa:牛生长激素聚腺苷酸化序列;orf3:插入位点下游的开放阅读框架3。orf69:orf70中插入位点上游的开放阅读框架69;p455:新颖的启动子;h3:转基因流感病毒血球凝集素;71pa:新颖的聚腺苷酸化序列;δorf70:含有orf71的启动子的orf70的其余部分,其编码结构性病毒糖蛋白ii(gpii)。图14.免疫印迹法:经rehv-1rach-se-1/3-p430-h1av-70-455-h3(b)、空载体rehv-1rach-se(se)感染或模拟感染(对照)的细胞的免疫印迹法。复制印迹用针对h3(h3)的商业兔高免疫血清(pa-34930)、针对h1(h1)的商业兔高免疫血清(pa-34929)或针对ehv-1gpii(gpii)的单克隆抗体ai2g7孵育。图15:经cav2cmviecpvvp2感染的ai-st2015细胞的流动式细胞测量分析:感染后72小时。图16:具有新颖的ehv-4启动子的rcav-2:经感染的ai-st2015细胞的流动式细胞测量分析:感染后48小时。图17:具有新颖的ehv-4启动子的rcav-2:经感染的e1bmdck(新颖的rcav-2)细胞中cpvvp2蛋白质表达的斑点印迹分析1°=1/50a-cpv-fitcmab(vmrd)2°=1/1.000山羊抗小鼠igg过氧化物酶(jir)a)原始斑点印迹数据b)由斑点印迹产生的半定量数据:对于定量,使用imagej软件(burger,w.,burge,m.j.(编),2008.digitalimageprocessing:analgorithmicintroductionusingjava.springer-verlag,newyork)分析点印迹。将图像颜色反转以减去所记录的各点的背景及积分密度。值如下分配+及-标示:“++++”=>800000,“+++”=500000至800000,“++”=300000至499999,“+”=120000至299999,“+/-”=80000至119999及“-”=<80000。图18:经rcav-2p455rabg感染的细胞中的rabg检测:在<1%的经原始rcav-2cmvierabg感染的细胞中检测到表达。图19:经rcav-2p455rabg感染的细胞中的rabg检测:a)经感染的ai-st2015细胞中cpvvp2表达的ifa,b)ai-st2015细胞中rabg表达的ifa,c)bivi2011mdck细胞中rabg表达的ifa-对rabg及cav-2进行双重染色。图20.各组在攻击前及攻击后1、2及3天的平均体温。误差条,标准偏差。按照研究日自左至右:阴性对照组(neg.ctrl.)、攻击对照组(chall.ctrl.)、经rach-se-70-p455-h3疫苗接种一次的动物(1×ehv-1)、经rach-se-70-p455-h3疫苗接种两次的动物(2×ehv-1)或经失活猪iav疫苗接种两次的动物(2×死疫苗)。图21.各组在攻击后一天及三天的平均肺得分。误差条,标准偏差。阴性对照组(neg.ctrl.)、攻击对照组(chall.ctrl.)、经rach-se-70-p455-h3疫苗接种一次的动物(1×ehv-1)、经rach-se-70-p455-h3疫苗接种两次的动物(2×ehv-1)或经失活猪iav疫苗接种两次的动物(2×死疫苗)。图22.在攻击日采集的动物血清对抗猪iavh3攻击病毒株r452-14的倒数血清中和(sn)效价。20,检测极限。阴性对照组(neg.ctrl.)、攻击对照组(chall.ctrl.)、经rach-se-70-p455-h3疫苗接种一次的动物(1×ehv-1)、经rach-se-70-p455-h3疫苗接种两次的动物(2×ehv-1)或经失活猪iav疫苗接种两次的动物(2×死疫苗)。图23.在猪iav攻击施加后一或两天获取的balf的il-1β结果。各点表示每一只动物所测定的值。阴性对照组(neg.ctr.)、攻击对照组(chall.ctr.)、经rach-se-70-p455-h3疫苗接种一次的动物(1×ehv-1)、经rach-se-70-p455-h3疫苗接种两次的动物(2×ehv-1)或经失活猪iav疫苗接种两次的动物(2×死疫苗)。图24.在第2次疫苗接种后7天再刺激的pbmc的ifnγ-elispot结果。(a)未经疫苗接种的对照组;(b)经失活猪iav疫苗疫苗接种两次;(c)经rehv-1rach-se-70-p455-h3疫苗接种一次;(d)经rehv-1rach-se-70-p455-h3疫苗接种两次。对于仅经rehv-1rach-se-70-p455-h3疫苗接种一次的动物,再刺激对应于第1次疫苗接种后7天。各点表示在给定时间点及在用特定刺激物再刺激后每一只动物所测定的值。对于再刺激,使用对应于rehv-1rach-se-70-p455-h3中的h3疫苗抗原重组表达的猪iavha(ha_v)、对应于攻击病毒株r452-14的h3重组表达的猪iavha(ha_ch)、稀释ha_v及ha_ch的培养基(rpmi)、空ehv-1载体rach-se(ehv-1空)、疫苗rach-se-70-p455-h3(ehv-1-h3)、猪iavh3n2攻击病毒株r452-14(h3n2)、用于生长r452-14的来自未感染细胞的细胞上清液(mdck)或重组表达的猪iav核蛋白(np)。图25.转移质粒pu1/3-p430-h1hu-bghkbgh的示意性图谱。图26:转移质粒pu70-p455-h1pdm-71k71的示意性图谱。图27:放大orf1/3及orf70插入区的rehv-1rach-se-1/3-p430-h1hu-70-p455-h1pdm(rehv-1rach-se_d)的线性双链dna基因组。图28:经rehv-1rach-se_b、rach-se_d、rach-se感染或未感染(对照)的细胞的免疫印迹。复制印迹用针对h3的多克隆兔高免疫血清(pa5-34930)、针对h1的多克隆兔高免疫血清(pa5-34929)或对抗ehv-1糖蛋白ii(gpii)的单克隆抗体(ai2g7)孵育。所有抗体产生预期的图案,证实所需抗原h3及h1的表达且不同病毒的复制效率相当,如由所有经感染细胞样品中ehv-1gpii的极类似的染色所判断。图29:小鼠血清的a型流感病毒中和测试的结果。*误差条指示标准偏差。图30:转移质粒pu70-455-sbvgc_71k71的图谱。图31:a)用于检测sbv病毒基因组的未经疫苗接种的对照牛(上图)及经rehv-sbv-gc疫苗接种两次的动物(下图)的定量rt-pcr的结果。对于每组未经疫苗接种及经疫苗接种的动物,分别由不同类型的线及符号来标识个别动物。动物1以具有黑色实心圆的黑线描绘(对应于图27b中的黑色条)。动物2以具有灰色实心三角形的灰色虚线描绘(对应于图27b中的浅灰色条)。动物3以具有空心方形的虚线描绘(对应于图27b中的白色条)。动物4以具有灰色实心菱形的灰色虚线描绘(对应于图27b中的深灰色条)。b)未经疫苗接种的对照牛(上图)及经rehv-sbv-gc疫苗接种两次的动物(下图)的血清中和测试结果。对于每组未经疫苗接种及经疫苗接种的动物,分别由不同条颜色/填充(自黑色经浅灰色及深灰色至白色)来标识个别动物。动物1以黑色条描绘(对应于图27a中具有黑色实心圆的黑线)。动物2以浅灰色条描绘(对应于图27a中具有灰色实心三角形的灰色虚线)。动物3以白色条描绘(对应于图27a中具有空心方形的黑色虚线)。动物4以深灰色条描绘(对应于图27a中具有灰色实心菱形的灰色虚线)。图32:ehv中和测试。自相应组中同一动物的样品获得的所有结果以相同的灰色色调展示:一个动物由黑色填充条表示、另一个动物由浅灰色填充条表示、第三个动物由白色条表示且第四个动物由深灰色条表示。图33:以在攻击后一天杀死的动物的每g肺组织的tcid50确定猪iav肺效价。neg.ctrl.,阴性对照组;chall.ctrl.,攻击对照组;2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。数据点指示个别动物所获得的平均值。中间水平线分别指示组平均值。上部及下部水平线分别指示标准偏差。各组成对统计学比较的p值在下方给出且通过分别使用曼-惠特尼检验(mann-whitneytest)及使用标准软件设置的windows软件graphpad7.02,graphpadsoftware,inc.,lajolla,ca92037,usa的t检验计算。图34:以在攻击后三天杀死的动物的每g肺组织的tcid50确定猪iav肺效价。neg.ctrl.,阴性对照组;chall.ctrl.,攻击对照组;2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。数据点指示个别动物所获得的平均值。中间水平线分别指示组平均值。上部及下部水平线分别指示标准偏差。各组成对统计学比较的p值在下方给出且通过分别使用曼-惠特尼检验及使用标准软件设置的windows软件graphpad7.02,graphpadsoftware,inc.,lajolla,ca92037,usa的t检验计算。图35:以在攻击后五天杀死的动物的每g肺组织的tcid50确定猪iav肺效价。neg.ctrl.,阴性对照组;chall.ctrl.,攻击对照组;2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。数据点指示个别动物所获得的平均值。中间水平线分别指示组平均值。上部及下部水平线分别指示标准偏差。各组成对统计学比较的p值在下方给出且通过分别使用曼-惠特尼检验及使用标准软件设置的windows软件graphpad7.02,graphpadsoftware,inc.,lajolla,ca92037,usa的t检验计算。图36:对针对重组表达的猪iav血球凝集素h3抗原的猪免疫球蛋白g(igg)具有特异性的酶联免疫吸附分析(elisa)结果,该抗原与由疫苗株rehv-1rach-se_b表达的h3同源。对于测试,各孔用100ng重组表达的h3涂布。分别成对测量样品,由成对测量值计算样品平均值,且由样品平均值计算组值。chall.ctrl.,攻击对照组(充当阴性对照物);2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。误差条指示标准偏差。研究天数(sd)以图例形式指示于图形右侧。图37:对针对重组表达的猪iav血球凝集素h3抗原的猪免疫球蛋白g(igg)具有特异性的酶联免疫吸附分析(elisa)结果,该抗原与由疫苗株rehv-1rach-se_b表达的h3同源。对于测试,各孔用100ng重组表达的h3涂布。分别成对测量样品,由成对测量值计算样品平均值,且由样品平均值计算组值。chall.ctrl.,攻击对照组(充当阴性对照物);2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。误差条指示标准偏差。研究天数(sd)以图例形式指示于图形右侧。图38:干扰素γ特异性酶联免疫吸附斑点分析(ifnγelispot)的结果。在研究第28天(sd28)自取自于研究动物的血液纯化外周血液单核细胞(pbmc)。pbmc随后用感染复数为1的h3n2猪iav攻击病毒株r452-14(h3n2moi1)或用浓度为1μg/ml的与由疫苗株rehv-1rach-se_b表达的h3同源的重组表达的猪iavh3抗原(rh31μg/ml)再刺激。使用经再刺激的pbmc,进行干扰素γ特异性酶联免疫吸附斑点分析(ifnγelispot),且分别将所获得的值相对于106个细胞标准化且计算作为每组的平均值。chall.ctrl.,攻击对照组(充当阴性对照物);2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。误差条指示标准偏差。图39:干扰素γ特异性酶联免疫吸附斑点分析(ifnγelispot)的结果。在研究第28天(sd28)自取自于研究动物的血液纯化外周血液单核细胞(pbmc)。pbmc随后用感染复数为1的h3n2猪iav攻击病毒株r452-14(h3n2moi1)或用浓度为1μg/ml的与由疫苗株rehv-1rach-se_b表达的h3同源的重组表达的猪iavh3抗原(rh31μg/ml)再刺激。使用经再刺激的pbmc,进行干扰素γ特异性酶联免疫吸附斑点分析(ifnγelispot),且分别将所获得的值相对于10^6个细胞标准化且计算作为每组的平均值。chall.ctrl.,攻击对照组(充当阴性对照物);2×im,肌肉内经疫苗接种两次的组;in+im,第一次鼻内及第二次肌肉内经疫苗接种的组;2×in,鼻内经疫苗接种两次的组。误差条指示标准偏差。实施例包括以下实施例以说明本发明的优选实施例。本领域技术人员应了解,以下实施例中所揭示的技术代表本发明人发现在本发明实施中起良好作用的技术,且因此可视为构成其优选实施方式。然而,根据本发明,本领域技术人员应了解,在不背离本发明的精神及范畴的情况下可对所揭示的特定实施例作出许多改变且仍获得相同或类似结果。实施例1:新颖的启动子的鉴别及构建鉴别适合的启动子序列的策略如下:ehv-4序列中在两个已知orf上游的600bp片段首先通过使其与ehv-1基因组的相应序列片段比对来进行分析。所选基因为编码主要衣壳蛋白(mcp)的orf42及编码糖蛋白g(gg)的orf70。主要衣壳蛋白为病毒粒子中最丰富的成分之一,且一旦新合成的病毒dna准备进行包装,就需要在细胞核中组装衣壳。因此,预期其启动子在病毒复制周期的早期及晚期具有活性。对于糖蛋白g,已知其基因(orf70)亦在复制周期的早期及晚期具有活性(colle等人1995,drummer等人1998)。假定的mcp启动子的序列相同性为82.2%且假定的gg启动子的序列相同性为82.3%。这些差异视为一方面足够大以防止同源重组,且另一方面足够小以允许在ehv-1复制期间转录活化。为测试启动子活性,合成600bpdna片段4pgg600及4pmcp600且克隆于编码自发荧光蛋白质mcherry的报告子基因的上游(shaner等人,2004)。作为转录终止信号及mrna稳定作用,将牛生长激素聚腺苷酸化序列(bghpa;goodwin及rottman,1992)直接克隆于报告子基因3'端下游。为用作阳性对照,cmv启动子自市售质粒pcdna3.1(invitrogen)扩增且克隆于mcherry报告子基因的上游,此处亦在报告子基因的3'端添加bghpa。用三种质粒(pblu-4pggmcherry、pblu-4pmcpmcherry及pblu-cmvmcherry)转染细胞培养物且通过荧光显微镜检查mcherry荧光。在转染后的不同时间,cmv启动子的强活性为明显的。4pgg600启动子在转染后亦具有活性,4pmcp600启动子的活性亦可检测,但与4pgg600启动子相比较弱且甚至在转染后三天当与cmv启动子相比时甚至弱得多。为研究病毒基因产物对启动子活性的效应,经pblu-4pgg600-mcherry或pblu-4pmcp600-mcherry转染的细胞培养物在转染后一天用绿色荧光ehv-1rachi-ef重复感染。病毒基因产物明显转活化4pmcp600启动子,与不存在ehv-1rachi-ef复制的情况相比,活性显著较高。该效应亦存在于用pblu-4pgg600-mcherry转染且用ehv-1rachi-ef重复感染的细胞培养物中,但并不如此剧烈,因为在不存在病毒复制的情况下的初始活性高于针对pblu-4pmcp600-mcherry所观察到的活性。尽管如此,对于两种600bp启动子,仍证明病毒复制对其在细胞培养物中的活性的转活化效应。若该等600bp序列含有通常位于活化子组件上游的抑制子组件,则可解释此效应。因此,在不存在病毒基因产物的情况下,较短启动子可更具活性。为测试此种情况,将两种ehv-4启动子序列截短至其原始长度的约75%且再次测试。具体而言,4pgg的600bp启动子截短至430bp,新名称:p430:tctatttgaggacccgccgagtaccccacaagagtatgtaaaaagctgtcattctcaactactgagaataatatcaaagctaaagataaaccctgaggagtttccacgggaaccagagtctaggctcgtgcgcggatacatcgaatacgccagcctagagcgtaagccacatacgcgctatccttgcttccagcgcgtgaacctacacattgacggggaatttttgatccataaaatgctagcgttcaatgctgcgatgcgcccatccgcagaagagttgttgtcctacccaatgtttatgaatctgtaggatgactaacagatttggggtggagacggcgtgggcgatactgtataaagttgtactacttaccagcccagtcagtgtgctgtagtgccaccacctgtaaagctgtgataagctgcagtt(seqidno:3)4pmcp截短至449bp,新名称:p455:ttggtggtagcatatactaccttatttatacgctccgagctgtttttcagcatgctagcacccaacgccgagcgagagtatataactcccatcattgcccacaagcttatgccacttattagcgtccgctctgccgtttgcttagtcataatatctaccgccgtttacgcagcagacgctatctgcgacacaattggatttgcgataccgcgcatgtggatgtgtattttaatgagatcaacctccatgaagcgtaactagggggcctcccactgaggcactaccggcttagcagctgactaacacagtataaaacgtgagaagaaatcagtctcatgcgccattagcgctaggctagttagcgtggaggaccggagcgctaccgccagcagtttcatccgcctggttacgggtttgttaacacctaccggtgttttaccgctaccata(seqidno:4)。将含有缩短的启动子的mcherry报告子质粒转染于细胞培养物中且通过荧光显微镜检查。虽然p430活性与600bp型式(4pgg600)相当,但p455的活性相比4pmcp600的活性显著增加。此结果与使用两种启动子的600bp型式的转染/重复感染实验的结果一致,即在同一细胞中ehv-1复制的存在提供4pmcp600启动子转活化的机制,强烈地增加其活性,而4pgg600启动子的转活化为可见的,但不太明显。除两种新颖的启动子之外,亦需要新颖的聚腺苷酸序列以便自新颖的orf70插入位点表达。该组件称为71pa。将其核苷酸序列合成且克隆于含有靶向prach-se中的orf70插入位点的p455的转移质粒中的mcherryorf的下游。接着,生成rehv-1rach-se以分析启动子在病毒复制背景中的活性(表1)。两种ehv-4启动子(p430及p455)、cmv启动子及小鼠细胞巨大病毒ie1启动子(mcmv)与bgh聚腺苷酸信号组合用于引导mcherry的表达以提高mrna稳定性。将如由等人(1985)所述的mcmvie1启动子(增强子)合成且克隆于质粒载体中,由此将其亚克隆于转移质粒中。另外,p455亦克隆于orf70中新颖的插入位点中,从而与新颖的聚腺苷酸信号71pa组合驱动mcherry表达。rehv-1rachmc70作为另一个对照物包括于实验中。经此重组病毒感染的细胞在内源性gg启动子(eggp)控制下表达mcherry(表1)。表1vero或pk/wrl细胞经感染复数为1的全部六种mcherry表达病毒感染。在感染后0、4、8及12小时采集经感染细胞且制备总rna。通过dna酶i消化破坏污染rna制备物的病毒及细胞基因组dna。在添加及不添加反转录酶的情况下通过反转录显示rna的完整性及病毒dna的移除,随后用特异性针对编码ehv-1的必需结构性糖蛋白d的orf72的引物对(第1130/1131号引物,(tgtctaccttcaagcttatg(seqidno:5)/ctagcgcagtcgcgttg(seqidno:6))进行pcr。预期的196bppcr产物仅由已添加反转录酶的反转录样品(cdna)扩增,具体而言,在t1=4hp.i.、t2=8hp.i.及t3=12hp.i.制备的样品,而非在t0=0hp.i.制备的样品。正如所预期,反转录酶尚未添加至反应中的所有样品均未产生任何pcr产物。因此,显示将用作qpcr模板的样品(cdna)不含病毒基因组dna。随后,使用特异性针对mcherry的引物对(第1079/1080号引物,(gcgaggaggataacatgg(seqidno:7)/acccttggtcaccttcag(seqidno:8))及orf72引物对1130/1131(tgtctaccttcaagcttatg(seqidno:5)/ctagcgcagtcgcgttg(seqidno:6)),通过qpcr分析用所添加的酶自反转录获得的cdna。orf72qpcr的ct值用于评定并行运行不同病毒感染的可比性且使mcherryqpcr的ct值标准化。因此,相对于感染后的时间及不同病毒定量mcherry的转录(图3)。如图3左图所示,六种不同病毒的orf72转录物的ct值在四个不同的感染后的时间几乎一致。理想地,全部六种病毒将在所研究的时间产生一致值且将仅可见一条线。几乎一致的线证实足够的实验质量,同样12hp.i.时间结果为有效的,因为与8hp.i.相比降低指示转录物数量的进一步增加,其仅当复制尚未达到其最大值时才可能。计算各感染后时间的统计平均值。各病毒在特定时间的值除以该时间所计算的平均值且用作将mcherryqpcr的ct值标准化以使其可直接比较的因子。mcherryqpcr的标准化ct值图解展示于图3右图中。线发散表明在不同的病毒感染细胞中产生的mcherry转录物数目的差异。在不同类型的图形中,组合两个实验,一个使用vero-eu细胞(v)且一个使用pk/wrl细胞(p)(图4)。如上所述,通过在反转录酶存在及不存在下反转录,随后用orf72引物进行pcr来确认rna制备物的质量及随时间推移的病毒复制。如上所述,基于orf72的qpcrct值使所获得的mcherry的qpcrct值标准化。t1=4hp.i.、t2=8hp.i.及t3=12hp.i.的标准化ct值自t0的标准化ct值减去(δ标准化ct),使得与转录活性的正相关。虽然在vero(v)或pk/wrl(p)细胞中的两个实验无法直接比较,但pk/wrl细胞中的较高表达量最可能反映pk/wrl对ehv-1复制的优越的容许性,其通常使得感染性病毒的效价高十倍。虽然源于ehv的启动子p430、p455及eggp的活性在所用细胞株的相应感染后时间几乎相同,与其插入位点或所用聚腺苷酸(bgh或71pa)无关,但cmv及mcmv启动子在pk/wrl细胞中的活性较高。在vero-eu细胞中,仅mcmv启动子显示具有较高活性,cmv启动子不优于ehv启动子。由这些实验得出结论,ehv-4启动子p430及p455适用于ehv-1rach主链中以驱动自orf1/3及orf70插入位点插入的转基因的表达。实施例2:在重组ehv-1载体疫苗中使用新颖的p455启动子及构建重组病毒p455启动子:对于第一动物实验,使用来自猪源性a型流感病毒的流感血球凝集素亚型h3(a/猪/意大利/7680/2001(h3n2),genbank登陆编号:abs50302.2)。合成其编码序列且亚克隆以产生转移载体pu70-p455-h3-71k71,将h3置于新颖的p455启动子及新颖的71pa聚腺苷酸化信号控制下且使盒与插入至orf70中的重组区成框(图5)。通过使用red重组系统顺道诱变(tischer等人2006),将表达盒p455-h3-71插入于prach-se的orf70中以生成prach-se70-p455-h3(图6)。pk/wrl细胞用prach-se70-p455-h3转染,拯救重组病毒rehv-1rach-se70-p455-h3且进行空斑纯化两次。表达盒的正确插入通过测序插入区的高保真度pcr产物来验证。通过间接免疫荧光分析(ifa,图7)分析经感染细胞中转基因的表达。使用单克隆抗体ai2g7(由bi拥有)通过ifa(未展示)及免疫印迹法(图8)确认编码ehv-1gpii的orf71的修复。经感染细胞的质膜上h3三聚体的出现通过使用鸡红血球的血球吸附测试(未展示)来分析。在pk/wrl细胞中以tcid50/ml形式确定的峰效价与亲本病毒rehv-1rach-se在相同的范围内,其表明转基因表达对病毒复制无不利效应(未展示)。此通过在拯救后rehv-1rach-se70-p455-h3在pk/wrl细胞中传代直至第20代(p20)来证实。在p5、p10、p15及p20通过滴定、测序及免疫印迹法(图8),在p10及p20另外通过ifa表征病毒,且确认ha编码插入物连同启动子及聚腺苷酸序列的ha表达及基因稳定性。图8中所示的两种印迹为用特异性检测ehv-1糖蛋白ii(gpii)的单克隆抗体ai2g7(左)或用来自饲养兔的对抗流感血球凝集素亚型h3的商业多克隆抗体(pa5-34930)(右)孵育的拷贝。正如所预期,在所有经重组ehv-1感染的细胞培养物中检测到gpii。正如所预期,在所有经rehv-1rach-se-70-p455-h3的不同传代感染的细胞中检测到全长h3。h3抗血清的特异性展示于同一免疫印迹中,参见条带gg430mc。正如所预期,此处仅gpiimab产生反应,而抗h3抗体未结合于相应的复本条带中。通过使用单克隆抗h3抗体及马抗ehv抗血清在经p20感染的细胞中的病毒空斑双重免疫荧光分析(difa),证实几乎所有诱生空斑的ehv-1亦表达h3(未展示)。所有测试均证实重组ehv-1rach-se-70-p455-h3的稳定性。实施例3:使用p455启动子的概念验证动物研究(poci)及血清学反应的评定测试动物:纳入准则及实验设计:如表2中所汇总,poc-i研究中包括五组由未处理的a型流感母猪生产的十只猪崽。表21×107tcid50感染剂量的rehv-1rach-70-p455-h3(ehv-1)在五周龄施用一次或在两周龄及五周龄施用两次。为进行比较,市售失活疫苗(inact)在两周龄及五周龄施用两次。所有猪崽均不含母系来源的抗体,以免消除失活疫苗(inact)的效应。两组未经疫苗接种但接受氯化钠生理溶液(nacl)注射,以分别充当攻击对照物或严格的阴性对照物。在第二次疫苗接种后21天,除严格的阴性对照组之外的所有组均用1×107tcid50异源a型流感(iva)病毒株(来自猪的h3n2a型流感病毒r452-14,由bi拥有的攻击分离株)攻击。虽然在未经疫苗接种的攻击对照组(challctrl)中,所有猪在攻击感染后一天及三天均在其肺中具有高流感病毒效价,但严格的阴性对照组(negctrl)及已经rehv-1rach-se-70-p455-h3接种两次的组(ehv2×)中的所有猪在两天均对iva呈阴性。在经失活对照疫苗接种两次的组(inact2×)中,五只动物中之一者在攻击后第三天具有低iva效价。在攻击前21天经rehv-1rach-se-70-455-h3接种一次的组(ehv1×)中,五只动物中的两者在攻击感染后一天且五只中之一者在攻击后三天在其肺中具有低iva效价。(图9)。经1×107tcid50rehv-1rach-se-70-p455-h3接种两次完全保护猪免受异源iav亚型h3n2的攻击感染。已证明ehv-1载体rach-se适于猪接种且新颖的启动子455在经疫苗接种的猪中具有驱动iav血球凝集素的免疫原性表达的功能。实施例4:在重组ehv-1载体疫苗中使用新颖的p430启动子及构建重组病毒p430启动子:新鉴别的p430启动子用于驱动来自h1n1病毒的另一流感血球凝集素((a/猪/gent/132/2005(h1n1),genbank登陆编号:afr76623.1)的表达。由于此病毒分离株中的血球凝集素基因来源于禽类iav,故其将称为h1av。合成h1av且亚克隆于orf1/3插入区的转移载体中以生成pu1/3-p430-h1av-bgh_k_bgh。h1av的表达在p430启动子及牛生长激素(bgh)聚腺苷酸信号控制下(图10)。通过使用red重组系统顺道诱变(tischer等人2006),将表达盒p430-h1av-bgh插入于prach-se的orf1/3中以生成prach-se1/3-p430-h1av(图11)。pk/wrl细胞用prach-se1/3-p430-h1av转染,拯救重组病毒rehv-1rach-se1/3-p430-h1av且进行空斑纯化两次。表达盒的正确插入通过测序插入区的高保真度pcr产物来验证。转基因在经感染细胞中的表达是使用市售单克隆及多克隆抗体通过间接免疫荧光分析(ifa)及免疫印迹法来分析(图12)。使用单克隆抗体ai2g7(由bi拥有)通过ifa及免疫印迹法确认编码ehv-1gpii的orf71的修复(未展示)。通过使用鸡红血球的血球吸附测试(未展示)分析h1av的正确加工及转运以及在经感染细胞的质膜中的定位。在pk/wrl细胞中以tcid50/ml形式确定的峰效价与亲本病毒rach-se在相同的范围内,其表明转基因表达对病毒复制无不利效应(未展示)。由抗体pa-34929在75kda迁移的宽带的特异性检测与由序列所预测的重组ha糖蛋白的预期外观一致。细胞膜用单克隆抗体c102的表观染色与所预期的亚细胞定位一致(图12)。为测试所表达的重组血球凝集素是否如所预期般的加工及转运,vero细胞经感染复数为0.01的rehv-1rach-se-1/3-p430-h1av、rehv-1rach-se-70-p455-h3、rehv-1rach-se(亲本)感染或保持未感染。24hp.i.经感染及未感染的活细胞用鸡红血球于pbs中的悬浮液培育,用pbs洗涤且用荧光hoechst33342细胞核染色剂染色。由于鸟类红血球含有细胞核,故其可用hoechst33342染色且通过荧光显微镜呈现为微小蓝色斑点,与经不表达血球凝集素的rehv-1rach-se感染的细胞相比,经rehv-1rach-se-1/3-p430-h1av或rehv-1rach-se-70-p455-h3感染的细胞上鸡红血球的吸附量显著增加(未展示)。由此可推断,血球凝集素以如同其是由真正的流感病毒感染产生般的方式翻译、加工及转运至经载体病毒感染的细胞的质膜。经感染细胞的血球吸附的明显表型支持免疫印迹法及免疫荧光分析的发现,显示转基因蛋白质的高效表达且表明在经ehv-1载体感染的细胞的细胞表面上形成功能性ha三聚体。实施例5:在重组ehv-1载体疫苗中在两个插入位点并行使用两种新颖的启动子p455及p430为展示两种新颖的启动子可并行使用,产生表达两种不同a型流感病毒亚型的两种不同血球凝集素的重组ehv-1rach。通过用单插入病毒rehv-1rach-se-70-p455-h3及rehv-1rach-se-1/3-p430-h1av感染的经感染细胞的免疫印迹验证多克隆商业抗体对h3(pa5-34930)及h1(pa5-34929)的特异性及缺乏交叉反应性(未展示)。以重组bacprach-se-70-p455-h3起始,通过两步red重组将在转移载体pu1/3-p430-h1av-bgh_k_bgh(图10)中组装的表达盒p430-h1av-bgh插入至orf1/3插入位点中以产生prach-se-1/3-p430-h1av-70-p455-h3。pk/wrl细胞用prach-se1/3-p430-h1av-70-p455-h3转染,拯救重组病毒rehv-1rach-se1/3-p430-h1av-70-p455-h3且进行空斑纯化两次(图13)。此重组病毒的简称为rehv-1rach-se_b。表达盒的正确插入通过测序插入区连同侧接序列的高保真度pcr产物来验证。转基因在经感染细胞中的表达是使用市售单克隆及多克隆抗体通过间接免疫荧光分析(ifa,未展示)及免疫印迹法来分析(图14)。编码ehv-1gpii的orf71的修复是使用单克隆抗体ai2g7(由bi拥有)通过ifa(未展示)及免疫印迹法来确认(图14)。如图14中所示,转基因h3及h1av在经双插入重组rehv-1rach-se-1/3-p430-h1av-70-p455-h3(b)感染的细胞培养物中并行表达。转基因表达为稳定的且未削弱测试的病毒效价,直至pk/wrl细胞中的第11代(未展示)。两种新颖的启动子p430及p455经展示在细胞培养物中的rehv1-rach复制的情形下具有功能性。在病毒复制周期期间的活性水平似乎与由活体外启动子动力学实验所推断的非常相似。这些特性允许基于ehv-1rach或以相似效率并行表达两种不同抗原的其他载体平台形成重组载体疫苗。若疫苗靶标由两种不同病原体组成,则两种新颖的启动子与两个聚腺苷酸化序列组合应用在两个插入位点中可显著降低货品的成本且呈现优于仅表达一种抗原成分的载体的明显优势。实施例6:使用新颖的p455及p430启动子的rcav-2载体疫苗方法ai-st2015细胞用以下rcav-2感染:1.cav-2cmviebrsv(抗cav2的阳性对照物;抗vp2的阴性对照物)2.cav-2p430cpvvp2(despliceda1-2-1)3.cav-2p430cpvvp2(gen0.95d1-5-1)4.cav-2p455cpvvp2(gen0.95e1-8-1)5.cav-2p430rabg(n)6.cav-2p455rabg(n)免疫荧光分析(ifa)ai-st2015细胞在感染后72小时用cytofix/cytoperm固定且用抗cpvvp2-fitc(mab)、抗rabg-fitc(mab)及抗cav-2-fitc(猪抗血清)(vmrd)染色。流式细胞测量术(fc)ai-st2015细胞在感染后48及72小时用cytofix/cytoperm固定且细胞用抗cav-2-fitc、抗cpvvp2-fitc(vmrd)、对抗cpv的猪高免疫血清(benchmark)及抗rabg-fitc(novus)染色。cpvvp2的斑点印迹来自经感染的e1bmdck(针对rcav-2)细胞的澄清(6000×g,5min)组织培养物上清液/溶解物(冷冻/融化)用pbs连续稀释,随后添加至设备中且经由抽吸吸附至pvdf。后续步骤为暴露于含5.0%biorad印迹级阻断剂的tbst30分钟,暴露于1°抗体1.0小时,三次tbst洗涤及暴露于过氧化物酶结合的2°抗体(抗小鼠及抗猪,jacksonimmunoresearch)1.0小时且经由tmb观测。对于定量,使用imagej软件(burger,w.,burge,m.j.(编),2008.digitalimageprocessing:analgorithmicintroductionusingjava.springer-verlag,newyork)分析点印迹。将图像颜色反转以减去所记录的各点的背景及积分密度。值如下分配+及-标示:“++++”=>800000,“+++”=500000至800000,“++”=300000至499999,“+”=120000至299999,“+/-”=80000至119999及“-”=<80000。cav-2vp2构建体:通过经rcav-2疫苗病毒感染的细胞产生病毒样颗粒(vlp)可为犬腺病毒(cav-2)疫苗功效的关键因素。虽然可拯救含有cmvie驱动的cpvvp2表达盒的rcav-2,但使用常规cmvie启动子无法在经rcav-2cmviecpvvp2感染的细胞中实现大量vp2表达(用于产生vlp)。无法拯救含有cmv5启动子的rcav-2vp2病毒。采用ifa、流式细胞测量术及斑点印迹评定经rcav-2感染的ai-st2015细胞中ehv-4启动子驱动的cpvvp2表达。用与cav-2fitc结合的猪多克隆抗体(vmrd)探测cav-2蛋白质表达。用小鼠单克隆(vmrd)及猪高免疫血清(benchmark)探测cpvvp2蛋白质表达。cav-2及cpvvp2蛋白质易于通过ifa观测且通过fc在相当大比例的经携带cpvvp2的两种不同核苷酸变体(despl及gen0.95,在感染后48及72小时)的rcav-2感染的ai-st2015细胞中检测到。通过斑点印迹在组织培养物上清液/溶解物(在冷冻/融化后)中鉴别出大量cpvvp2蛋白质(且极有可能反映组装的vlp的存在)。在经感染的ai-st2015细胞中易于通过ifa检测到cpvvp2表达(参见图19a)。在小于3%的经原始rcav-2cmviecpvvp2感染的细胞中检测到cpvvp2表达(参见图15)。因此,拯救携带由新颖的源于ehv4的启动子p430及p455驱动的cpvvp2表达盒的rcav-2且测试以确定其是否可能有效驱动cpvvp2在经感染细胞中的表达。出人意料地,在14%至36%的经感染细胞中检测到cpvvp2表达(参见图16)。另外,与原始rcav-2cmviecpvvp2(其中斑点印迹分析展示cmvie驱动的cpvvp2信号处于或低于背景水平-与来自经cav-2、rcav-2cmviebrsvf感染的细胞的上清液/溶解物及来自未感染细胞的细胞培养物上清液/溶解物相当-数据未展示)相比,在来自经感染的ai-st2015细胞的上清液/溶解物中检测到大量cpvvp2(参见图17)。图17展示vp2蛋白质可在上清液中识别到且因此预期呈所需构象(vlp)以具有免疫原性。重要的是,当cag或cmv5启动子序列存在于位于e3区中的表达盒中时,未实现重组cav-2的拯救。其似乎具有序列特异性,因为表达盒的尺寸尚未超过所观察到实验基因组尺寸限制。因此,本发明的新颖的源于ehv-4的启动子序列,诸如p430及p455,不仅有助于转基因表达,且亦支持病毒拯救的关键步骤。总之,ifa、fc及斑点印迹证明在经感染的aist2015细胞中,稳固ehv-4启动子驱动由rcav-2表达cpvvp2转基因。这些结果证实ehv-4启动子在除ehv-1以外的载体中的效用。rcav-2rabg(n)构建体:使用本发明的新颖的源于ehv-4的p455启动子生成第二cav-2构建体。选择rcav-2rabg(n)是因为使用常规cmvie启动子未观察到经感染细胞的表达。此实验的目标为通过测量经rcav-2p455rabg(n)感染的ai-st2015细胞中ehv-4启动子驱动的rabg蛋白质表达来证实在rcav-2具有第二转基因rabg(膜蛋白)的情形下新颖的ehv-4启动子的活性。采用ifa及流式细胞测量术评定经rcav-2感染的ai-st2015细胞中ehv-4启动子驱动的rabg表达。用与cav-2fitc结合的猪多克隆抗体(vmrd)探测cav-2蛋白质表达。用小鼠单克隆抗体(novus)探测rabg蛋白质表达。cav-2及rabg蛋白质易于通过ifa观测且通过fc在经携带rabg(n)的rcav-2感染的ai-st2015细胞中检测到(在感染后72小时)。因此:虽然rabg易于在经rcav-2p455rabg感染的细胞中检测到(参见图19b及c),但在<2.0%的经原始rcav-2cmvierabg感染的细胞中检测到表达(参见图18)。在经具有不相关转基因的rcav-2感染的细胞中发现极少乃至无信号(参见图19b,cpvvp2,其与抗rabg抗体不相关)。如双重染色的图19c中所见,许多cav-2阳性细胞亦为rabg阳性。总的,ifa及fc数据证明ehv-4启动子驱动经感染的aist细胞通过rcav-2表达rabg转基因。这些结果进一步证实本发明的ehv-4启动子在除ehv-1以外的载体中的效用。实施例7:猪的单价ehv-1载体a型流感病毒疫苗(h3疫苗)的产生、活体外表征及活体内测试选择血清型h3的猪iav流感病毒血球凝集素(seqidno27)(a/猪/意大利/7680/2001(h3n2),genbank登陆编号:abs50302.2)作为待测试抗原用于猪的疫苗接种研究。此种对抗猪iav的新疫苗提供diva特征,例如通过在经猪iav野毒株感染的动物中而非在仅经本文所述的疫苗接种的动物中检测对抗猪iav蛋白质np或na的抗体,因为其仅表达一种猪iavha蛋白质。合成其编码序列且亚克隆以产生转移载体pu70-p455-h3-71k71,将h3置于新颖的p455启动子及新颖的71pa聚腺苷酸化信号控制下且使盒与插入至orf70中的重组区成框(图1及5)。通过使用red重组系统顺道诱变,将表达盒p455-h3-71插入于prach-se的orf70中以产生prach-se70-p455-h3。pk/wrl细胞用prach-se70-p455-h3转染,拯救重组病毒rehv-1rach-se70-p455-h3且进行空斑纯化两次。(图6)。表达盒的正确插入通过测序插入区的高保真度pcr产物来验证。转基因在经感染细胞中的表达是使用市售单克隆及多克隆抗体通过间接免疫荧光分析(ifa,图7)及免疫印迹法(图8)来分析。使用单克隆抗体ai2g7(由bi拥有)通过ifa(未展示)及免疫印迹法(图8)确认编码ehv-1gpii的orf71的修复。经感染细胞的质膜上h3三聚体的出现通过使用鸡红血球的血球吸附测试(未展示)来分析。在pk/wrl细胞中以tcid50/ml形式确定的峰效价与亲本病毒rach-se在相同的范围内,其表明转基因表达对病毒复制无不利效应(未展示)。此通过在拯救后rehv-1rach-se70-p455-h3在pk/wrl细胞中传代直至第20代(p20)来证实。在p5、p10、p15及p20通过滴定、测序及免疫印迹法(图8),在p10及p20另外通过ifa表征病毒,且确认ha编码插入物连同启动子及聚腺苷酸序列的ha表达及基因稳定性。图8中所示的两种印迹为用特异性检测ehv-1糖蛋白ii(gpii)的单克隆抗体ai2g7(左)或用来自饲养兔的对抗流感血球凝集素亚型h3的商业多克隆抗体(pa5-34930)(右)培育的复本。正如所预期,在所有经重组ehv-1感染的细胞培养物中检测到gpii。正如所预期,在所有经rehv-1rach-se-70-p455-h3的不同传代感染的细胞中检测到全长h3。h3抗血清的特异性亦通过用其他表达来自h1亚型病毒的流感血球凝集素的重组ehv-1rach-se感染的细胞的免疫印迹来展示,图14。通过使用单克隆抗h3抗体及马抗ehv抗血清在经p20感染的细胞中的病毒空斑双重免疫荧光分析(difa),证实几乎所有诱生空斑的ehv-1亦表达h3(未展示)。所有测试均证实重组ehv-1rach-se-70-p455-h3的稳定性。为研究作为载体疫苗在猪崽中的特性,在疫苗接种-攻击研究中测试rehv-1rach-se-70-p455-h3。具体地,无对抗猪iav的母系来源免疫性(无母本抗体)的猪崽在两周龄及五周龄(二次注射接种,2×ehv-1)或仅在五周龄(一次注射接种,1×ehv-1)肌肉内经剂量为1×107tcid50的rach-se-70-p455-h3疫苗接种两次。未经疫苗接种的组充当阴性对照物且根据制造商说明书(除疫苗接种时间点之外)在两周龄及五周龄经市售失活猪iav疫苗疫苗接种的一组动物充当阳性对照物(杀死)。在8周龄,除阴性对照物外的所有动物受气管内施用的1×107tcid50剂量的h3n2猪iav攻击病毒株(欧洲野生病毒分离株r452-14,其h3与rach-se-70-p455-h3中所用的h3疫苗抗原异源)攻击。未经疫苗接种且未经攻击的动物充当阴性对照物,而未经疫苗接种但经攻击的动物充当攻击对照物。在疫苗接种时及疫苗接种后以及在攻击之前及之前,测量体温且在不同时间点获取血液样品。在攻击后一天,杀死每组一半的动物且针对猪iav感染的典型病变对肺进行评分,每只动物分别取出每一左肺及右肺三个肺样品以确定肺匀浆中的感染性猪iav效价,且对支气管肺泡灌洗液(balf)进行取样。在攻击后三天对每组剩余的一半动物执行相同程序。当研究猪iav攻击病毒施用后的体温升高时,未经疫苗接种的动物在攻击后1天显示体温升高约1℃。对于经rach-se-70-p455-h3疫苗接种两次的组,在攻击后1天的此体温升高得以避免(图20)。在猪iav攻击病毒施用后1或3天杀死的动物的肺得分的评定揭露阴性对照物未显示猪iav感染的典型肺病变,攻击对照物显示在6-7%的平均范围的肺病变,而关于组平均值,经rach-se-70-p455-h3疫苗接种两次的组的肺病变得分剧烈降低至1至小于4%(图21)。在猪iav攻击病毒施用后1或3天杀死的动物的平均猪iav肺效价显示阴性对照物在肺样品中未显示猪iav,而攻击对照物显示每g肺组织的病毒效价在大于5(第3天)至大于7log(第1天)范围内。形成鲜明对比的是,经rach-se-70-p455-h3疫苗接种一次的组的组平均值剧烈降低至约2log或更小,且经rach-se-70-p455-h3疫苗接种两次的组降低至不可检测的水平(图22)。当在疫苗接种后测试猪iav中和抗体的诱导时,经rach-se-70-p455-h3疫苗接种一次的动物的血清在第一次疫苗接种后三周显示在约160范围内的倒数中和效价,且经rach-se-70-p455-h3疫苗接种两次的动物的血清在第2次疫苗接种后三周显示约2560的中和效价,而未经疫苗接种的组的血清不具有可检测的猪iav中和抗体水平(图22)。当在猪iav攻击后1或3天测定动物的balf中促炎性细胞介素il-1β的量时,在第1天测试的四只动物中的三只可检测到大于100pg/ml直至900pg/ml的il-1β水平,而对于经rach-se-70-p455-h3疫苗接种一次的动物的balf,这些水平降低至100-300pg/mlil-1β,且所有经rach-se-70-p455-h3疫苗接种两次的动物的水平甚至进一步降低至0至小于100pg/mlil-1β(图23)。此显示经rach-se-70-p455-h3疫苗接种已有效预防在猪iav感染后诱导促炎性细胞介素il-1β。当测试在研究第28天取样的外周血液单核细胞(pbmc)的再刺激且使用不同刺激物时,刺激来自未经疫苗接种的动物的pbmc在ifnγ-elispot中显示小于75/1×106个计数,与所用刺激无关(图24a)。已接受失活疫苗两次的动物(杀死)的pbmc在用重组猪iav核蛋白np再刺激时显示约150/1×106个计数且在用猪iavh3n2攻击病毒株r452-14再刺激时在ifnγ-elispot中显示约3000/1×106个计数,但当使用重组猪iavha或ehv-1病毒时不显示pbmc的再刺激(75/1×106个计数或更低的水平)(图24b)。相比的下,经rach-se-70-p455-h3疫苗接种一次或两次的动物在用猪iavh3n2攻击病毒株r452-14再刺激时亦在ifnγ-elispot中显示约200(1×ehv-1)至300(2×ehv-1)/1×106个计数,但当使用重组猪iavnp时无pbmc再刺激(75/1×106个计数或更低的水平)(图24c及d)。当ehv-1病毒用于再刺激时,经rach-se-70-p455-h3疫苗接种一次或两次的动物在用空ehv-1疫苗rach-se再刺激时在ifnγ-elispot中显示约300/1×106个计数,且当分别使用表达猪iavh3的rach-se-70-p455-h3疫苗时,此值进一步增加至大于400/1×106个计数(图24c及d)。因此,当重组猪iavha用于再刺激时,仅经rach-se-70-p455-h3疫苗接种一次或两次的动物在ifnγ-elispot中显示约100-150(1×ehv-1)至150-200(2×ehv-1)/1×106个计数(图24c及d)。实施例8猪的四价ehv-1载体a型流感病毒疫苗的产生、活体外表征及活体内测试如下所述,在所述发明中,如所述来源于h1n2、h3n2、h1n1禽类及h1n1流行性猪iav亚型/血清型的四种猪iav血球凝集素(ha)抗原是由两种重组ehv-1载体病毒表达。此种对抗猪iav的新四价疫苗提供diva特征,例如通过在经猪iav野毒株感染的动物中而非在仅经本文所述的疫苗接种的动物中检测对抗猪iav蛋白质np或na的抗体,因为其仅表达猪iavha蛋白质。新四价猪iav疫苗活体外表征且活体内测试其对抗猪iav的功效。新鉴别的p430启动子用于驱动猪iavh1n1((a/猪/gent/132/2005(h1n1),genbank登陆编号afr76623.1)的表达。由于此病毒分离株中的血球凝集素基因来源于禽类iav,故其将称为h1av。合成h1av且亚克隆于orf1/3插入区的转移载体中以生成pu1/3-p430-h1av-bghkbgh。将h1av的表达置于p430启动子及牛生长激素(bgh)聚腺苷酸信号的控制下且与插入至orf1/3中的重组区成框(图10)。通过使用red重组系统顺道诱变,将表达盒p430-h1av-bgh插入于prach-se的orf1/3中以产生prach-se1/3-p430-h1av)。pk/wrl细胞用prach-se1/3-p430-h1av转染,拯救重组病毒rehv-1rach-se1/3-p430-h1av图11且进行空斑纯化两次。表达盒的正确插入通过测序插入区的高保真度pcr产物来验证。转基因在经感染细胞中的表达是使用市售单克隆及多克隆抗体通过间接免疫荧光分析(ifa)及免疫印迹法来分析(图12)。使用单克隆抗体ai2g7(由bi拥有)通过ifa及免疫印迹法确认编码ehv-1gpii的orf71的修复(未展示)。通过使用鸡红血球的血球吸附测试(未展示)分析h1av的正确加工及转运以及在经感染细胞的质膜中的定位。在pk/wrl细胞中以tcid50/ml形式确定的峰效价与亲本病毒rach-se在相同的范围内,其表明转基因表达对病毒复制无不利效应(未展示)。由抗体pa-34929在75kda迁移的宽带的特异性检测与由序列所预测的重组ha糖蛋白的预期外观一致。细胞膜用单克隆抗体c102的表观染色与所预期的亚细胞定位一致。为测试所表达的重组血球凝集素是否如所预期般的加工及转运,vero细胞经感染复数为0.01的rehv-1rach-se-1/3-p430-h1av、rehv-1rach-se-70-p455-h3、rehv-1rach-se(亲本)感染或保持未感染。24hp.i.经感染及未感染的活细胞用鸡红血球于pbs中的悬浮液培育,用pbs洗涤且用荧光hoechst33342细胞核染色剂染色。由于鸟类红血球含有细胞核,故其可用hoechst33342染色且通过荧光显微镜呈现为微小蓝色斑点,与经不表达血球凝集素的rehv-1rach-se感染的细胞相比,经rehv-1rach-se-1/3-p430-h1av或rehv-1rach-se-70-p455-h3感染的细胞上鸡红血球的吸附量显著增加(未展示)。由此可推断,血球凝集素以如同其是由真正的流感病毒复制产生般的方式翻译、加工及转运至经载体病毒感染的细胞的质膜。经感染细胞的血球吸附的表型支持免疫印迹法及免疫荧光分析的发现(关于h1av,图12),显示转基因蛋白质的高效表达且表明在经ehv-1载体感染的细胞的细胞表面上形成功能性ha三聚体。通过用单插入病毒rehv-1rach-se-70-p455-h3及rehv-1rach-se-1/3-p430-h1av感染的经感染细胞的免疫印迹验证多克隆商业抗体对h3(pa5-34930)及h1(pa5-34929)的特异性及缺乏交叉反应性(未展示)。接着,生成表达两种不同a型流感病毒亚型/血清型的两种不同血球凝集素的重组ehv-1rach-se。以重组bacprach-se-70-p455-h3起始,通过两步red重组将在转移载体pu1/3-p430-h1av-bgh_k_bgh(图10)中组装的表达盒p430-h1av-bgh插入至orf1/3插入位点中以产生prach-se-1/3-p430-h1av-70-p455-h3。pk/wrl细胞用prach-se1/3-p430-h1av-70-p455-h3转染,拯救重组病毒rehv-1rach-se1/3-p430-h1av-70-p455-h3且进行空斑纯化两次。此重组病毒的简称为rehv-1rach-se_b(图13)。表达盒的正确插入通过测序插入区连同侧接序列的高保真度pcr产物来验证。转基因在经感染细胞中的表达是使用市售单克隆及多克隆抗体通过间接免疫荧光分析(ifa,未展示)及免疫印迹法来分析(图14)。编码ehv-1gpii的orf71的修复是使用单克隆抗体ai2g7(由bi拥有)通过ifa(未展示)及免疫印迹法来确认(图14)。转基因h3及h1av在经双插入重组rehv-1rach-se_b感染的细胞培养物中并行表达。转基因表达为稳定的且未削弱测试的病毒效价,直至pk/wrl细胞中的第11代。已显示具有两个插入位点及两个新颖的启动子的增强型ehv-1载体并行表达两种流感病毒血球凝集素。如通过ifa所测定的亚细胞定位及如通过免疫印迹法所测定的sds-page中的迁移率对应于自文献已知的经a型流感病毒感染的细胞中表达的真正的血球凝集素。接着,生成表达血球凝集素h1hu,seqidno:29,(a/猪/意大利/4675/2003(h1n2);genbank登陆编号adk98476.1)及h1pdm,seqidno:26,(a/猪/意大利/116114/2010(h1n2);genbank登陆编号adr01746.1)的第二双插入rehv-1rach。合成h1hu的编码序列且亚克隆于orf1/3插入区的转移载体中以产生pu1/3-p430-h1hu-bghkbgh。将h1hu的表达置于p430启动子及牛生长激素(bgh)聚腺苷酸信号的控制下且与插入至orf1/3中的重组区成框(图25)。合成h1pdm的编码序列且亚克隆以产生转移载体pu70-p455-h1pdm-71k71,将h1pdm置于新颖的p455启动子及新颖的71pa聚腺苷酸化信号控制下且使盒(cassette)与插入至orf70中的重组区成框(图26)。随后,通过使用red重组系统顺道诱变将表达盒p430-h1av-bgh及p455-h1pdm-71插入至prach-se中,首先生成prach-se-1/3-p430-h1hu。使用此经修饰的bac作为靶标,通过使用red重组系统顺道诱变插入p455-h1pdm-71,生成prach-se-1/3-p430-h1hu-70-p455-h1pdm。将prach-se-1/3-p430-h1hu-70-p455-h1pdm转染于pk/wrl细胞中,拯救rehv-1rach-se-1/3-p430-h1hu-70-p455-h1pdm且进行空斑纯化三次。新颖的重组载体病毒的简称为rehv-1rach-se_d(图27)。转基因在经感染细胞中的表达是使用市售单克隆及多克隆抗体通过间接免疫荧光分析(ifa,未示出)及免疫印迹法来分析(图25)。编码ehv-1gpii的orf71的修复是使用单克隆抗体ai2g7(由bi拥有)通过ifa(未展示)及免疫印迹法来确认(图28)。重组rehv-1的基因稳定性及表型稳定性通过在细胞培养物中传代,每5代确定病毒效价来展示。每十代确认插入区的序列以及通过免疫印迹法确认转基因表达(未展示)。表达保真度通过在甲基纤维素覆盖下空斑的双重ifa,计数用抗ehv抗体及转基因特异性抗体染色的空斑来评定(未展示)。为研究作为载体疫苗在猪崽中的特性,在疫苗接种-攻击研究中测试由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗。具体地,具有对抗猪iav的母系来源免疫性(母本抗体阳性)的猪崽在一周龄及四周龄(二次注射接种,2×ehv-1)或仅在四周龄(一次注射接种,1×ehv-1)肌肉内经剂量为每一疫苗病毒株1×107tcid50的rehv-1rach-se_b及rehv-1rach-se_d疫苗接种两次。未经疫苗接种的组充当阴性对照物。在11周龄,除阴性对照物外的所有动物受气管内施用的1×106tcid50剂量的h3n2猪iav攻击病毒株(欧洲野生病毒分离株r452-14,其h3与rehv-1rach-se_b中所用的h3疫苗抗原异源)攻击。未经疫苗接种且未经攻击的动物充当阴性对照物,而未经疫苗接种但经攻击的动物充当攻击对照物。在疫苗接种时及疫苗接种后以及在攻击之前及之前,测量体温且在不同时间点获取血液样品。在攻击后一天,杀死每组一半的动物且针对猪iav感染的典型病变对肺进行评分,每只动物分别取出每一左肺及右肺三个肺样品以确定肺匀浆中的感染性猪iav效价,且对支气管肺泡灌洗液(balf)进行取样。在攻击后三天对每组剩余的一半动物执行相同程序。分析样品材料及收集的数据以尤其确定攻击后的体温变化、猪iav感染后的临床征象、肺得分、猪iav肺效价、肺组织的组织学变化、猪iav血清中和效价、balf中的细胞介素水平、如通过ifnγ-elispot所测量的pbmc再刺激及b细胞活化。实施例9在经二价rehv-1rach疫苗接种的小鼠中诱导对抗两种抗原的中和抗体反应将rehv-1rachseb(rehv-1rach-se-1/3-p430-h1av-7-p455-h3,参见图13)用于balb/c小鼠的免疫以证明所表达的转基因在除猪之外的另一物种中具有免疫原性,且通过鼻内施用诱导对抗两种抗原中的任一者的中和抗体。具体地,将三组每组五只3-5周龄balb/c小鼠在研究第0天及第21天分别鼻内接种以40μlrehv-1rachseb(rehv-1rach-se-1/3-430-h1av-7-455-h3,第1组)或40μl空载体(rehv-1rach-se,第2组,载体对照物)或40μl组织培养基(第3组阴性对照物)。对于第1组及第2组,感染性重组ehv-1剂量分别为1×105tcid50/40μl。在研究第0天(在第1次接种前)、第7天、第14天、第21天(在第2次接种前)、第28天及第35天对小鼠进行抽血。由血液样品制备血清且在-80℃下冷冻储存。用于检测对抗载体病毒的抗体的免疫荧光分析在0.001的感染复数(moi)下,用自空载体bacprach-se1.2拯救的病毒rehv-1rach-se1212感染ai-st细胞。感染后24小时观察到独特空斑且加工细胞以用于间接免疫荧光分析(ifa)。测试以1:50稀释于pbs中的全部三组最终血液(在第二次疫苗接种后14天获得)的血清。作为阳性对照物,来自经ehv-1接种的马的血清以1:500的稀释度使用。二级抗体为市售fitc结合的兔抗小鼠igg用于小鼠血清及cy5结合的山羊抗马igg用于马血清且以1:200稀释度使用。通过荧光显微镜评估抗体结合。所有经疫苗接种的小鼠已产生在ifa中与经rehv-1rach-se感染的细胞反应的抗体。未感染的细胞未与测试血清中的任一者结合。来自阴性对照组小鼠的血清既未对经感染的细胞亦未对未感染的细胞显示任何特异性结合。数据汇总于下表中。表3.关于抗ehv-1抗体的ifa的荧光显微术结果由此可推断,rehv-1接种至小鼠鼻孔中引起感染及病毒复制,以便刺激小鼠免疫系统以产生抗ehv-1抗体。病毒中和测试(vnt)为显示对抗所表达的来源于a型流感病毒(iav)(a/猪/意大利/7680/2001(h3n2))或(a/猪/gent/132/2005(h1n1))的转基因的保护性免疫的诱导,测试小鼠血清对抗各个病毒的中和活性(allwinn等人2010;trombetta等人2014)。用于中和测试的iav为2014年来自德国的猪的分离株,具体而言a/猪/德国/ar452/2014(h3n2)及a/猪/德国/ar1181/2014(h1n1)。由于这些病毒株与疫苗靶标所来源的病毒株为异源的,故这些病毒通过小鼠血清的任何中和将表明通过rehv-1疫苗接种对保护性免疫的广泛且有效的诱导。使用已显示流感病毒抗体阴性的猪血清作为阴性对照血清。a型流感病毒中和测试:用于病毒中和以及在96孔培养盘中反滴定的mdck细胞在使用之前在37℃/5%co2下培育两天。将相应iav储备液h3n2及h1avn1在冰上融化且在含有庆大霉素(gentamycin)及双倍浓度的胰蛋白酶的mem(mem/genta/2×胰蛋白酶)中稀释。测试的血清来自第1组(rehv-1rachseb)、第2组(空载体)、阳性对照组(经失活多价iav疫苗接种的猪的血清及阴性对照组的最终血液。将血清加热失活且分别在两个及三个独立测试中,以1:16开始连续1:2稀释直至1:4096。iav稀释至约100tcid50/中和反应。中和反应物在37℃、5%co2下培育2小时。一式四份地对所用病毒进行反滴定。移除生长培养基且用含有庆大霉素及胰蛋白酶的培养基洗涤mdck细胞,随后添加中和反应物或反滴定的病毒稀释液。在将中和反应物或病毒稀释液添加至mdck细胞后,vnt及滴定盘分别在37℃/5%co2下培育1小时。此后,移除接种体且用含有庆大霉素及胰蛋白酶的新鲜培养基覆盖细胞。感染后五天,监测及记录cpe。测试中实际使用的病毒效价根据reed及münch以tcid50/ml形式计算,且报导测试血清预防流感病毒诱导典型cpe的稀释度,参见下表。表4:流感h1avn1vnt结果表5:流感h3n2vnt结果为比较独立测试的结果,通过将倒数血清稀释度与由其中和的相应效价相乘来计算中和能力。三个测试的平均值随后除以100以反映100tcid50的中和(表3、4及5)。数据汇总且图解展示于图29中。经rehv-1rachseb疫苗接种的所有小鼠已产生对抗相应iav(亚型h3n2及h1avn1的异源病毒株)的中和抗体。因此,两倍鼻内施用自p455启动子(h3)控制下的orf70插入位点及并行地自p430启动子(h1av)控制下的orf1/3插入位点表达iav的血球凝集素的rehv-1rach-se成功地刺激balb/c小鼠的保护性免疫反应。可得出结论,载体rehv-1rach-se可用于在鼻内疫苗接种后并行表达两种不同转基因以刺激免疫反应。实施例10牛的ehv-1载体化施马伦贝格(sbv)病毒疫苗的产生、活体外表征及活体内测试一种新出现的崩芽病毒(bunyavirus)为施马伦贝格病毒(schmallenbergvirus,sbv),其为第一种欧洲辛波血清群(simbuserogroup)病毒(正布尼亚病毒属(genusorthobunyavirus)),当怀孕的动物在妊娠的关键阶段感染时,可能导致流产、死产及严重的胎儿畸形,且其现在愈来愈多地用作研究正布尼亚病毒的模型病毒(bilk等人,2012)。由于辛波病毒通过昆虫媒介传播且无可用的治疗选择方案,故疫苗接种为疾病控制的主要组成部分。对抗sbv及其他辛波病毒(诸如赤羽病毒(akabanevirus,akav)或艾罗病毒(ainovirus))的失活全病毒疫苗为可用的,且已开发对抗sbv的活减毒疫苗(anonymous,2013,2015;kraatz等人,2015;wernike等人,2013b),然而,这些疫苗中无一者允许区分野外感染及经疫苗接种的动物(diva原则)。直到最近才在致死性小动物攻击模型及牛中测试基于来自sbv糖蛋白gc的氨基端的234个氨基酸(aa)的diva兼容性亚单位疫苗(wernike等人,2017)。当作为表达质粒递送或在哺乳动物细胞培养系统中表达时,gc结构域在至多66%的动物中赋予保护,而经连接于相关akav的相应结构域的sbv的gc结构域免疫的所有动物均受完全保护(wernike等人,2017)。为研究rehv-1rach-se作为载体疫苗在牛中的应用,将sbv-gc的234个氨基端aa插入至orf70(us4)插入位点中且在新颖的p455启动子及71pa聚腺苷酸信号控制下表达并在牛中的疫苗接种-攻击试验中进行测试。表达源于施马伦贝格病毒(sbv)糖蛋白c(gc)的抗原的重组ehv-1的产生施马伦贝格病毒(sbv)糖蛋白c(gc)的编码区的234个氨基酸部分经密码子使用优化以便在ehv-1中表达,且另外经修饰以实现有效转运及插入至经感染细胞的质膜中。为此,将来源于a型流感病毒(iav)血球凝集素(ha)亚型h1n2(a/猪/意大利/116114/2010(h1n2),genbank登陆编号adr01746.1)的信号肽编码序列以及来自ha的跨膜锚(tm)及细胞质c端分别附接至5'及3'端。另外,将gs连接子hmggsggggsggggsgggt(seqidno:30)插入于gc部分与ha-tm结构域之间。合成dna(seqidno:31)且亚克隆于pu70-455-71k71的noti/kpni位点中,pu70-455-71k71为通过red介导的bacprach-se重组将转基因表达盒插入至ehv-1的orf70(us4)中的转移载体。所得质粒pu70-455-sbvgc_71k71(图30)用xbai切割以释放3056bpdna片段(seqidno:32),将其转化至携带prach-se的大肠杆菌k12gs1783中。seqidno:31:包括用于亚克隆的限制性位点的合成dna序列seqidno:32:用于red重组以产生prach-se-70-455-sbvgc的dna片段通过星号(*)指示限制酶切割位置制备重组prach-se-70-455-sbvgcdna且通过使用herculasetm的高保真度pcr及pcr产物的sanger测序确认表达盒的正确插入及序列相同性。所用引物参见表6,seqidno:33至seqidno:37。表6:用于pcr及测序的引物#名称序列用途seqidno:33up70_f5'-cgtgcgcggatacatcg-3'pcr及测序seqidno:34up71_r5'-cgcttcgcaggtgggc-3'pcr及测序seqidno:35seq455-f15'-gactggtggtagcatatac-3'测序seqidno:36sbvgcf15'-gatcaacgagatcatctcc-3'测序seqidno:37sbvgcr15'-ctggagagagccgttgc-3'测序重组ehv-1rach-se-70-455-sbvgc的拯救及表征bacdna是由prach-se-70-455-sbvgc的四个不同纯系制备。将ai-st细胞(boehringer-ingelheim专有的猪睪丸细胞株)以105个细胞/孔的密度接种于6孔培养盘(corningincorporated-lifesciences,onebectoncircle,durham,nc27712,usa;ref353046)的含有10%fbs(sigma-aldrichchemiegmbh,munich,germany,safc,cat12003c-1000ml)的mem(sigma-aldrichchemiegmbh,munich,germany,safc62892-1000m3056)中。当细胞60-70%汇合时,通常次日,使用mirustmmrna转染试剂盒(mirusbiollc,545sciencedrive,madison,wi53711usa)根据供货商说明书用2μgbacdna转染细胞。简言的,将200μloptimemtm(thermofisherscientific)培养基添加至5ml聚苯乙烯管中。添加dna且混合。接着添加3μlboost试剂且通过涡漩混合,随后添加相同体积的转染试剂且再次通过涡漩混合。混合物在室温下培育3分钟且随后直接逐滴添加至细胞培养物中。细胞在37℃/5%co2下培育五天。将细胞冲洗至培养基中且采集以便在-80℃下储存。在mem中制备拯救病毒的1:10连续稀释液且涂覆于6孔培养盘中汇合的ai-st细胞单层上。在37℃/5%co2下吸附1小时后,移除接种体且用含有0.5%methocel(methylcelluloseph.eur.,fluka64632-500g)及5%fbs(mem-methocel)的半固体培养基覆盖细胞。在37℃/5%co2下培育两至三天(第1代)后,将距离相邻空斑尽可能远的个别空斑以10μl的体积抽吸且接种于6孔培养盘中的新ai-st细胞培养物中。培育经感染细胞两至三天,直至观察到大量cpe(第2代)。将细胞冲洗至培养基中且采集以便在-80℃下储存。重复此空斑纯化程序两次。加工经三次空斑纯化的病毒感染的ai-st细胞以分别用于间接免疫荧光分析(ifa)或免疫印迹法。将由经感染细胞制备的病毒dna用作使用herculasetm的高保真度pcr的模板。通过sanger测序分析所获得的pcr产物且确认具有理论序列的插入区与bac的相应pcr产物序列的相同性。间接免疫荧光分析用连续稀释于mem中的经三次空斑纯化的病毒感染24孔培养盘(corningincorporated-lifesciences,onebectoncircle,durham,nc27712,usa;ref353047)中的ai-st细胞。自细胞吸出生长培养基且用250μl经稀释的病毒(稀释度10-2至10-7)覆盖细胞。细胞在37℃/5%co2下培育1小时以吸附病毒,随后移除接种体且用1000μlmem-methocel/孔覆盖细胞并在37℃/5%co2下培育两天。当用显微镜观察到空斑形成时,细胞经加工以用于ifa。抽吸培养基且用1mlpbs(gibcolifetechnologies,paisleypa49rf,uk,dpbs(1x)ref14190-136)/孔洗涤细胞一次。移除pbs且通过添加1毫升/孔-20℃冷的乙醇(carlrothgmbh,schoemperlenstr.3-5,d-76185karlsruhe,art.nr.5054.1)固定细胞并在室温下培育30分钟。抽吸乙醇且风干细胞。细胞在室温下用1毫升/孔pbs复水10分钟后,添加稀释于pbs中的初级抗体(150微升/孔)且在室温下孵育1小时。移除初级抗体且用1mlpbs/孔洗涤细胞三次,持续2分钟,随后添加二级抗体稀释液(150微升/孔)。在室温下避光孵育1小时后,移除二级抗体稀释液且用1mlpbs/孔洗涤细胞三次,持续2分钟,且最后用500μlpbs/孔覆盖以通过荧光显微镜检查。所用抗体列在表7中。表7抗体稀释马抗ehv-1高免疫血清(boehringeringelheim兽医学研究中心专有)1:400抗sbv-gc单克隆抗体(wernike等人,2015a)1:50fitc结合的山羊抗小鼠iggjacksonimmunoresearch目录号115-095-0031:200cytm5结合的山羊抗马iggjacksonimmunoresearch目录号108-175-0031:200免疫印迹法1.感染:6孔培养盘中三个孔的汇合的ai-st细胞单层各通过分别将50μl及10μl融化的病毒储备液直接添加至生长培养基而用感染复数约为1的rehv-1rach-se-455-sbvgc的两种不同空斑分离株(#121.131p6及#121.232p6)及rehv-1rach-se1212p9的空斑分离株(自亲本空bacprach-se1.2拯救)感染。三个孔保持未感染。培育经感染及未感染的细胞两天且随后加工用于免疫印迹法。2.制备溶解物:补充有蛋白酶抑制剂混合液的ripa缓冲液(ripa+pi)制备如下:将0.7ml10×ripa溶解缓冲液millipore目录号20-188添加至6.3mlh2o,fisherscientific目录号bp2470-1中且将1片剂completetm微型蛋白酶抑制剂混合液(roche目录号11836153001)溶解于7ml1×ripa缓冲液中。将未感染的对照物刮入培养基中且将来自三个重复孔的悬浮液汇集在15ml离心管中并置于冰上。将经感染细胞冲洗入培养基中且将来自三个重复孔的悬浮液汇集在15ml离心管中并置于冰上。通过在4℃下以1000×g离心5分钟使细胞沉降。小心地抽吸上清液且使细胞集结粒再悬浮于ripa+pi中(未感染的细胞在300μl中,经感染细胞在150μl中)。悬浮液在冰上孵育30分钟且每10分钟进行涡旋。将悬浮液转移至1.5ml微量离心管中且通过在4℃下在微量离心机中以15000rpm离心10分钟而使未溶解的材料沉降。将澄清的上清液转移至新的1.5ml微量离心管中且储存在-80℃下直至使用。3.sds-page及在耐纶膜上转移:材料:bioradcriteriontgx无染色剂预制凝胶,4-20%,26孔,目录号_567-8095;bioradprecisionplus双色标记,目录号161-0374;bioradprecisionplus全蓝标记,目录号161-0373;bioradtransblotturbo转移试剂盒,midi格式目录号170-4159;biorad4×laemmli样品缓冲液(目录号161-0747)(bioradlaboratoriesgmbh,heidemannstrasse164,d-80939münchen);tgs操作缓冲液(sambrook等人),封闭溶液1:含5%fbs的pbst(sambrook等人);pbst。在不添加还原剂的情况下制备样品。样品在冰上融化且与1体积的4×缓冲液混合,在96℃下煮沸6分钟且保持在室温下直至凝胶装载。凝胶在230ma下操作30分钟且随后组装以使用bioradtransblotturbo系统电转移。转移设定成2.5a25v10分钟。膜在无菌蒸馏h2o中冲洗且在4℃下用25ml封闭溶液含5%fbs的pbst孵育30分钟。抗体孵育及检测材料:immun-starwesternc化学发光试剂盒(bioradlaboratoriesgmbh,heidemannstrasse164,d-80939münchen)目录号170-5070初级抗体:a:sbv-gc蛋白质特异性单克隆抗体(wernike等人,2015a)1:20b:针对ehv-1gpii的小鼠单克隆抗体ai2g7(boehringeringelheim专有)二级抗体:过氧化物酶结合的山羊抗小鼠,(jacksonimmuneresearch#115-035-146)1:5000所有孵育均在足够的体积中在恒定搅动下进行。将抗体稀释于5%fbs/tbst中。初级抗体在4℃下孵育隔夜。移除抗体溶液且用tbst洗涤印迹三次,持续5-10分钟。经稀释的二级抗体与印迹一起在室温下孵育1小时,移除且用tbst洗涤印迹三次,持续5-10分钟。将印迹置于透明的塑料薄片保护器上。1ml+1ml混合过氧化物及lumino/增强子溶液(各印迹总计2ml),用移液管移取于印迹上且孵育3至5分钟。此后,将膜置于chemidocxrs成像系统(bioradlaboratoriesgmbh,heidemannstrasse164,d-80939münchen)中且使用imagelab软件记录信号。病毒滴定在感染前一天,将ai-st细胞以2×104个细胞/孔接种于96孔盘(corningincorporated-lifesciences,onebectoncircle,durham,nc27712,usa;ref353072)的补充有10%fbs的mem中。将病毒储备液快速融化且置于冰上。在mem中制备10个连续1:10稀释液,每个稀释液1.2ml体积。将100微升/孔的病毒稀释液添加至细胞,每个稀释液一个垂直列的8个孔。各培养盘的垂直列11及12通过添加100微升/孔mem充当培养基对照物。一式三份地进行滴定且细胞在37℃/5%co2下培育5天。用显微镜检查细胞培养物且记录观察到ehv-1rach典型cpe的孔。效价根据reed及muench(1938)的方法以tcid50/ml形式计算。用于疫苗接种的重组ehv-1的表征通过免疫印迹法及双重免疫荧光分析(difa)显示经修饰的sbvgc234在经感染细胞中的表达,用于rehv-1rach-se-70-455-sbvgc121.232的空斑分离。使用多克隆马抗ehv抗血清及单克隆抗sbv抗体的difa证实转基因在明显100%的经rehv-1感染的细胞中的表达。当进行经rehv-1rach-se-70-455-sbvgc_121.232感染的细胞的difa时,用马抗ehv抗血清(紫色)染色的ehv-1抗原阳性细胞亦结合针对sbvgc的单克隆抗体。在非还原条件下进行的免疫印迹法证实经修饰的sbvgc234在经重组ehv-1rach-se-70-455-sbvgc感染的细胞中的表达。进行用针对sbvgc的单克隆抗体或针对ehv-1gpii的单克隆抗体探测经感染或未感染细胞的溶解物的免疫印迹法。虽然ehv-1gpii在所有经感染细胞中表达,但sbvgc仅在经rehv-1rach-se-70-455-sbvgc感染的细胞中,而不在经空载体rehv-1rach-se1212感染的细胞中表达。在模拟感染的细胞的溶解物中未检测到病毒蛋白质。用对抗ehv-1的gpii的单克隆抗体孵育平行印迹证实在转染后拯救重组病毒期间通过自我切除程序修复orf71(us5)。由经三次空斑纯化的分离株rehv-1rach-se-70-455-sbvgc_121.232培养的p7病毒原液在ai-st细胞中复制达到1.85×109tcid50/ml的极高效价,表明转基因的表达不会削弱此细胞株中的ehv复制。呈tcid50/ml形式的rehv-1rach-se-70-455-sbvgc_121.232的六次滴定的平均值得到1.85×109tcid50/ml,标准偏差为1.28×109tcid50/ml。动物及实验设计4只德国驯养品种的牛相隔三周经108tcid50rehv-sbv-gc疫苗接种两次;另4只牛保持作为未经疫苗接种的对照物。在第二次免疫后三周,所有动物皮下接种以2×0.5ml仅在牛中传代的sbv野毒株(wernike等人,2012)。在整个研究期间,每日测量直肠体温且由兽医检查动物的临床征象。每隔一周采集血清且通过市售基于n的elisa(id施马伦贝格病毒竞争,idvet,france)及通过如先前所述的针对sbv分离株bh80/11的微量中和测试(wernike等人,2013a)来分析。通过评定3天后的细胞病变效应进行评估;一式四份地测试所有样品且根据behrens及kaerber以nd50形式计算抗体效价。另外通过针对ehv病毒株rach(rehv-sbv-gc组及未经疫苗接种的对照动物)的微量中和测试分别分析在免疫日、攻击感染日及研究结束时采集的血清。在攻击感染后的前10天,在每日基础上另外采集血液样品。使用kingfisher96flex(thermoscientific,braunschweig,germany)与magattractvirusminim48试剂盒(qiagen,hilden,germany)的组合根据制造商说明书自这些样品提取病毒rna且通过基于s区段的实时rt-pcr进行测试(bilk等人,2012)。实验方案已由可靠的国家伦理委员会审查且经主管部门(德国罗斯托克梅克伦堡-前波莫瑞州农业食品安全及渔业局(stateofficeforagriculture,foodsafetyandfisheriesofmecklenburg-vorpommern,rostock,germany),参考lallfm-vtsd/7221.3-1.1-004/12)批准。临床观测结果及病毒rna检测在整个研究期间,无一只动物显示任何相关的sbv特异性临床征象且在直肠测量时,所有动物的体温保持在正常范围内。自攻击感染后第一天或第二天起,在各未经疫苗接种的对照动物的血清样品中连续四天可检测到病毒rna。在整个采样时段中,来自rehv-sbv-gc组的所有经疫苗接种的动物通过定量rt-pcr显示降低的病毒rna浓度(图31a)。在整个采样时段中,rehv-sbv-gc组的两只动物通过定量rt-pcr测试完全呈阴性(图31a)。在经rehv-sbv-gc免疫的两只动物中,分别以降低的水平检测到sbv基因组三天或五天。抗体反应在未经疫苗接种的对照动物中,在攻击感染之前,通过血清中和测试未检测到sbv特异性抗体。在感染后一或两周,在所有未经疫苗接种的动物中检测到高效价的中和抗体(图31b)。与未经疫苗接种的对照组相比,在经rehv-sbv-gc免疫的四头牛的两头中,在攻击感染当天可检测到sbv特异性中和抗体。在此组剩余的两只动物中,在攻击感染之前未检测到sbv特异性中和抗体,但在感染后两周存在中和抗体(图31b)。所有四只经疫苗接种的动物中sbv特异性中和抗体的效价均低于攻击对照物,表明攻击病毒的病毒复制效率低,且因此支持定量rt-pcr数据。ehv中和测试在mem中以1:5开始制备血清的两倍稀释液。将50μl含有100tcid50的sbv的mem及50μl稀释的血清在96孔细胞培养盘中孵育2小时。此后,添加100μl新鲜制备的bhk细胞(在含有10%胎牛血清的mem中)的悬浮液且培养盘在37℃/5%co2下孵育3-4天。通过光学显微镜评估细胞病变效应。所有血清均一式两份地测试,且根据behrens(个人通信)修改的kaerber(1931)以nd50形式计算抗体效价。如图32中所示的结果表明用rehv-1rach-se-70-455-sbvgc接种牛使得载体病毒的复制足够有效以诱导特异性免疫反应。在四只动物ehv-1中之一者中,在初次疫苗接种后三周可检测到极低效价的中和抗体(1:4)。在两次疫苗接种后,在第二次施用后三周,所有四头牛均已产生效价为1:128的中和抗体。由此结果可推断,ehv-1rach亦可具有作为牛的疫苗载体的功能。实施例11由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗对抗猪崽中的猪iavh3n2攻击的功效为研究在猪崽中作为载体化疫苗的特性,在第二项接种疫苗-攻击研究中测试由rehv-1rach-se_b(rehv-1rach-se-1/3-p430-h1av-70-p455-h3,参见图13)及rehv-1rach-se_d(rehv-1rach-se-1/3-p430-h1hu-70-p455-h1pdm,参见图27)组成的四价猪iav疫苗。在此第二项研究中,来自未经疫苗接种的母猪且在第一次疫苗接种时通过使用h3特异性elisa(图36)及通过病毒中和测试(数据未展示)针对猪iav特异性抗体测试呈血清学阴性的猪崽经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价疫苗接种两次。动物分别在其生命第一周(研究第0天,sd0)第一次及在其生命第四周(研究第21天,sd21)第二次以每一疫苗株、动物及疫苗接种2ml剂量中含1×107tcid50的剂量分别肌肉内且随后肌肉内(2×im)、或首先鼻内且随后肌肉内(in+im)、或两次鼻内(2×in)疫苗接种。未经疫苗接种的组充当阴性对照物且另一个未经疫苗接种的组充当攻击对照物。在其生命第十一周(研究第69或70天,sd42/43),除阴性对照物之外的所有动物均受2×107tcid50的h3n2猪iav攻击病毒株(欧洲野生病毒分离株r452-14,其h3与rehv-1rach-se_b中所用h3疫苗抗原异源)的气管内施用剂量攻击。未经疫苗接种且未经攻击的动物充当阴性对照物(neg.ctrl.),而未经疫苗接种但经攻击的动物充当攻击对照物。在疫苗接种时及疫苗接种后以及在攻击前,在不同时间点获取血液样品。在攻击后一天,杀死每组一半的动物且每只动物分别获取每个左肺及每个右肺三个肺样品。随后,测定每只动物每克肺组织匀浆的感染性猪iav效价作为每只动物左肺及右肺的平均值,其各由汇集的每个左肺或右肺三个样品的匀浆获得且分别相对于左肺或右肺的三个样品的总重量标准化。在攻击后三天对每组剩余的一半动物执行相同程序。对于所有疫苗接种组,自该组中的个别动物在攻击后第一天(ch+1)获取的样品所获得的感染性猪iav效价的中值在与攻击对照组相比时统计学上显著降低,而来自阴性对照组的所有动物均未在其肺匀浆中显示感染性猪iav病毒效价(图33)。此外,对于所有疫苗接种组,自该组中的个别动物在攻击后第3天(ch+3)获取的样品所获得的感染性猪iav效价的中值在与攻击对照组相比时统计学上显著降低,而来自阴性对照组的所有动物均未在其肺匀浆中显示感染性猪iav病毒效价(图34)。因此,经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗疫苗接种分别统计学上显著降低猪崽在用异源猪iavh3n2病毒株攻击后一天及三天的猪iav肺负荷。因此,本文所述的疫苗有效对抗猪体内的猪iav。此外,在研究第0天(sd0,在第一次疫苗接种前)、在研究第21天(sd21,在第二次接种疫苗前)及在研究第42或43天(sd42/43,在施用攻击材料前)取自于研究动物的血清通过对针对重组表达的猪iavh3抗原的免疫球蛋白g(igg)具有特异性的酶联免疫吸附分析(elisa)来分析,该重组表达的猪iavh3抗原与由疫苗株rehv-1rach-se_b表达的h3同源。虽然来自阴性对照组的血清的平均od值仅给出所有测量时间点的极低值,但来自疫苗接种组的血清在两次肌肉内施用(2×im;sd21及sd42/43)后、在第一次鼻内且随后肌肉内施用(in+im;sd42/43)后及在两次鼻内施用(2×in;sd42/43)后表达出强烈的od值增加;图36。因此,经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗疫苗接种分别在猪崽体内引发针对由疫苗株rehv-1rach-se_b表达的猪iav血球凝集素h3的血清学免疫反应。另外,在研究第28天(sd28)自取自于研究动物的血液纯化外周血液单核细胞(pbmc)。pbmc随后用感染复数为1的h3n2猪iav攻击病毒株r452-14(h3n2moi1)或用浓度为1μg/ml的与由疫苗株rehv-1rach-se_b表达的h3同源的重组表达的猪iavh3抗原(rh31μg/ml)再刺激。使用经再刺激的pbmc,进行干扰素γ特异性酶联免疫吸附斑点分析(ifnγelispot),且分别将所获得的值相对于106个细胞标准化且计算作为每组的平均值(图38)。虽然来自攻击对照组(充当此测试的阴性对照物,动物未经疫苗接种)的经再刺激的pbmc在任一次再刺激后显示每组的平均斑点低于45个,来自经疫苗接种的动物的经再刺激的pbmc在任一次再刺激后分别显示在两次肌肉内施用后每组平均斑点超过85个,在第一次鼻内且随后肌肉内施用(in+im)后超过100个斑点及在两次鼻内施用(2×in)后超过150个斑点(图38)。因此,经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗接种在猪崽体内分别引发对抗由疫苗株rehv-1rach-se_b表达的猪iav血球凝集素h3及对抗用于异源攻击病毒感染的猪iavh3n2r452-14的细胞免疫反应。因此,猪崽经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗接种在猪崽体内诱导可检测的血清学及细胞免疫反应,且通过统计学上显著降低在异源猪iav攻击后一天及三天肺匀浆中的猪iav负荷来证明疫苗功效。实施例12由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗对抗具有母系来源抗体的猪崽中的猪iavh3n2攻击的功效为研究作为载体疫苗在猪崽中的特性,在第三项疫苗接种-攻击研究中测试由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗。在此第三项研究中,使用由在妊娠期间经针对猪iav的市售失活疫苗接种两次的母猪生产且喂养初乳及乳汁的猪崽。在第一次疫苗接种经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价疫苗接种两次时,通过使用h3特异性elisa(图37)及通过使用市售猪iav特异性抗体elisa(idexxinfluenzaaidexx,westbrook,maine04092,usa)按照制造商的测试建议对猪崽的猪iav特异性抗体血清学测试呈阳性(数据未展示)。动物分别在其生命第一周(研究第0天,sd0)第一次及在其生命第四周(研究第21天,sd21)第二次以每一疫苗株、动物及疫苗接种2ml剂量中含1×107tcid50的剂量分别肌肉内且随后肌肉内(2×im)、或首先鼻内且随后肌肉内(in+im)、或两次鼻内(2×in)疫苗接种。未经疫苗接种的组充当阴性对照物且另一个未经疫苗接种的组充当攻击对照物。在其生命第十一周(研究第69或70天,sd69/70),除阴性对照物之外的所有动物均受2×107tcid50的h3n2猪iav攻击病毒株(欧洲野生病毒分离株r452-14,其h3与rehv-1rach-se_b中所用h3疫苗抗原异源)的气管内施用剂量攻击。未经疫苗接种且未经攻击的动物充当阴性对照物,而未经疫苗接种但经攻击的动物充当攻击对照物。在疫苗接种时及疫苗接种后以及在攻击前,在不同时间点获取血液样品。在攻击后五天,杀死动物且每只动物分别获取每个左肺及每个右肺三个肺样品。随后,测定每只动物每克肺组织匀浆的感染性猪iav效价作为每只动物左肺及右肺的平均值,其各由汇集的每个左肺或右肺三个样品的匀浆获得且分别相对于左肺或右肺的三个样品的总重量标准化。对于所有疫苗接种组,自该组中的个别动物在攻击后第五天(ch+5)获取的样品所获得的感染性猪iav效价的中值在与攻击对照组相比时统计学上显著降低,而来自阴性对照组的所有动物均未在其肺匀浆中显示感染性猪iav病毒效价(图35)。因此,经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗疫苗接种分别统计学上显著降低猪崽在用异源猪iavh3n2病毒株攻击后五天的猪iav肺负荷。因此,本文所述的疫苗有效对抗猪体内的猪iav。此外,在研究第0天(sd0,在第一次疫苗接种前)、在研究第21天(sd21,在第二次接种疫苗前)及在研究第35天(sd35,在第二次接种疫苗后两周)取自于研究动物的血清通过对针对重组表达的猪iavh3抗原的免疫球蛋白g(igg)具有特异性的酶联免疫吸附分析(elisa)来分析,该重组表达的猪iavh3抗原与由疫苗株rehv-1rach-se_b表达的h3同源。虽然来自阴性对照组的血清的平均od值仅给出sd21及sd35的极低值,但来自疫苗接种组的血清在两次肌肉内施用(2×im;sd35)后、在第一次鼻内且随后肌肉内施用(in+im;sd35)后及在两次鼻内施用(2×in;sd35)后表达出强烈的od值增加;图37。因此,经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗疫苗接种分别在猪崽体内引发针对由疫苗株rehv-1rach-se_b表达的猪iav血球凝集素h3的血清学免疫反应。另外,在研究第28天(sd28)自取自于研究动物的血液纯化外周血液单核细胞(pbmc)。pbmc随后用感染复数为1的h3n2猪iav攻击病毒株r452-14(h3n2moi1)或用浓度为1μg/ml的与由疫苗株rehv-1rach-se_b表达的h3同源的重组表达的猪iavh3抗原(rh31μg/ml)再刺激。使用经再刺激的pbmc,进行干扰素γ特异性酶联免疫吸附斑点分析(ifnγelispot),且分别将所获得的值相对于106个细胞标准化且计算作为每组的平均值(图39)。虽然来自攻击对照组(充当此测试的阴性对照物,动物未经疫苗接种)的经再刺激的pbmc在任一次再刺激后显示每组的平均斑点低于15个,来自经疫苗接种的动物的经再刺激的pbmc在任一次再刺激后分别显示在两次肌肉内施用后每组平均斑点超过30个,在第一次鼻内且随后肌肉内施用(in+im)后超过55个斑点及在两次鼻内施用(2×in)后超过65个斑点(图39)。因此,经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗接种在猪崽体内分别引发对抗由疫苗株rehv-1rach-se_b表达的猪iav血球凝集素h3及对抗用于异源攻击病毒感染的猪iavh3n2r452-14的细胞免疫反应。因此,猪崽经由rehv-1rach-se_b及rehv-1rach-se_d组成的四价猪iav疫苗接种在猪崽体内诱导可检测的血清学及细胞免疫反应,且通过统计学上显著降低在异源猪iav攻击后五天肺匀浆中的猪iav负荷来证明疫苗功效。本文中所揭示及主张的所有组合物及方法均可根据本发明制备及执行,无需过多实验。虽然已根据优选实施例描述本发明的组合物及方法,但本领域技术人员应清楚变化可在不背离本发明的概念、精神及范畴的情况下施加于本文所述的组合物及方法及方法的步骤或步骤顺序。更具体地,显而易知在化学上及生理上相关的某些药剂可取代本文所描述的药剂,同时获得相同或类似结果。本领域技术人员显而易见的所有此类类似取代及修改视为在由随附申请专利范围所界定的本发明的精神、范畴及概念内。参考文献以下参考文献特定地以引用的方式并入本文中,以致其对本文所阐述的那些提供例示性程序或其他细节补充。1.allwinnr,geilerj,bergera,cinatlj,doerrhw.2010.determinationofserumantibodiesagainstswine-origininfluenzaavirush1n1/09byimmunofluorescence,haemagglutinationinhibition,andbyneutralizationtests:howistheprevalencerateofprotectingantibodiesinhumans?medmicrobiolimmunol.199(2):117-21.doi:10.1007/s00430-010-0143-4.epub2010feb17.2.anonymous(2013).vmdauthorizessbvvaccineforuseintheuk.theveterinaryrecord172,5433.anonymous(2015).schmallenbergvirusvaccine.theveterinaryrecord177,3214.bilks,schulzec,fischerm,beerm,hlinaka,hoffmannb(2012).organdistributionofschmallenbergvirusrnainmalformednewborns.veterinarymicrobiology159,236-2385.boshartm,weberf,jahng,k,fleckensteinb,schaffnerw.1985.averystrongenhancerislocatedupstreamofanimmediateearlygeneofhumancytomegalovirus.cell41(2):521-30.6.bustin,s.2000.absolutequantificationofmrnausingreal-timereversetranscriptionpolymerasechainreactionassays.journalofmolecularendocrinology25(2):169-193.7.charoensawan,v.,wilson,d.,teichmann,s.a.2010.genomicrepertoiresofdna-bindingtranscriptionfactorsacrossthetreeoflife.nucleicacidsres.38(21):7364-778.colle,c.f.3rd,o'callaghan,d.j.1995.transcriptionalanalysesoftheuniqueshortsegmentofehv-1strainkentuckya.virusgenes;9(3):257-68.9.k.,keil,g.m.,weber,f.,jasin,m.schaffner,w.,andkoszinowski,u.h.1985.alongandcomplexenhanceractivatestranscriptionofthegenecodingforthehighlyabundantimmediateearlymrnainmurinecytomegalovirus.pnasvol.82:8325-8329.10.drummer,h.e.,studdert,m.j.,crabb,b.s.1998.equineherpesvirus-4glycoproteingissecretedasadisulphide-linkedhomodimerandispresentastwohomodimericspeciesinthevirion.j.gen.virol.79:1205-121311.fields,b,knipe,d.m.;andhowley,p.m.2013.virology.6thed.philadelphia;wolterskluwerhealth/lippincottwilliams&wilkins12.foecking,m.k.,hofstetter,h.1986.powerfulandversatileenhancer-promoterunitformammalianexpressionvectors.gene45(1):101-5.13.goodwin,e.c.&rottman,f.m.1992.the3'flankingsequenceofthebovinegrowthhormonegenecontainsnovelelementsrequiredforefficientandaccuratepolyadenylation.j.biol.chem.267:16330-16334.14.hübert,p.h.,birkenmaier,s.,rziha,h.-j.andosterrieder,n.(1996),alterationsintheequineherpesvirustype-1(ehv-1)strainrachduringattenuation.journalofveterinarymedicine,seriesb,43:1-14.doi:10.1111/j.1439-0450.1996.tb00282.x15.g(1931)beitragzurkollektivenbehandlungpharmakologischerreihenversuche.archivfexperimentpatholupharmakol.;162:480-48316.kim,d.w.,uetsuki,t.,kaziro,y.,yamaguchi,n.,sugano,s.1990.useofthehumanelongationfactor1alphapromoterasaversatileandefficientexpressionsystem.gene16;91(2):217-23.17.kraatzf,wernikek,hechingers,p,granzowh,reimanni,beer,m(2015).deletionmutantsofschmallenbergvirusareavirulentandprotectfromviruschallenge.jvirol89,1825-183718.luke,gaandryan,md.2013.theproteincoexpressionprobleminbiotechnologyandbiomedicine:virus2aand2a-likesequencesprovideasolution.futurevirology,vol.8,no.10,pages983-996.19.ma,g.,eschbaumer,m.,said,a.,hoffmann,b.,beer,m.,osterrieder,n.2012.anequineherpesvirustype1(ehv-1)expressingvp2andvp5ofserotype8bluetonguevirus(btv-8)inducesprotectioninamurineinfectionmodel.plosone.2012;7(4):e34425.doi:10.1371/journal.pone.0034425.epub2012apr12.20.ma,g.,azab,w.,osterrieder,n.2013.equineherpesvirusestype1(ehv-1)and4(ehv-4)--mastersofco-evolutionandaconstantthreattoequidsandbeyond.vetmicrobiol.167(1-2):123-34.21.nolan,t.rebeccaehands,r.e.,andbustins.a.2006.quantificationofmrnausingreal-timert-pcrnatureprotocols1:1559-158222.osterrieder,n.,neubauer,a.,brandmüller,c.,kaaden,o.r.,ando'callaghan,d.j.1996.theequineherpesvirus1ir6proteininfluencesvirusgrowthatelevatedtemperatureandisamajordeterminantofvirulence.virology226:243-251.23.ptashne,m.2014.thechemistryofregulationofgenesandotherthingsthejournalofbiologicalchemistryvol.289,(9)5417-5435.reed,l.j.,andmuench,h.1938.asimplemethodofestimatingfiftypercentendpoints.am.j.hyg.(27)3;493-497.24.reedljandmuenchh(1938).asimplemethodestimatingfiftypercentendpoints.theamericanjournalofhygiene27(3)493-49725.rosas,c.t.,konig,p.,beer,m.,dubovi,e.j.,tischer,b.k.,osterrieder,n.,2007a.evaluationofthevaccinepotentialofanequineherpesvirustype1vectorexpressingbovineviraldiarrheavirusstructuralproteins.j.gen.virol.88(3),748-757.26.rosas,c.t.,b.k.tischer,g.a.perkins,b.wagner,l.b.goodman,n.osterrieder.2007b.live-attenuatedrecombinantequineherpesvirustype1(ehv-1)inducesaneutralizingantibodyresponseagainstwestnilevirus(wnv)virusresearch,125,pp.69-78.27.rosas,c.t.,vandewalle,g.r.,metzger,s.m.,loelzer,k.,dubovi,e.j.,kim,s.g.,parrish,c.r.,osterrieder,n.,2008.evaluationofavectoredequineherpesvirustype1(ehv-1)vaccineexpressingh3haemagglutininintheprotectionofdogsagainstcanineinfluenza.vaccine26(19),2335-3234.28.said,a.,elkelange,e.,beer,m.damiani,a.,osterrieder,n.2013.recombinantequineherpesvirus1(ehv-1)vaccineprotectspigsagainstchallengewithinfluenzaa(h1n1)pmd09virusresearch173:371-37629.sambrookjandrusselldw(2001).molecularcloning,3rded.coldspringharborlaboratorypress,coldspringharbor,newyork;isbn978-087969-577-430.shaner,n.c.,campbell,r.e.,steinbach,p.a.,giepmans,b.n.,palmer,a.e.,tsien,r,y.2004.improvedmonomericred,orangeandyellowfluorescentproteinsderivedfromdiscosomasp.redfluorescentprotein.natbiotechnol.dec;22(12):1567-72.epub2004nov21.31.tischer,b.k.,voneinem,j.,kaufer,b.,osterrieder,n.,2006.two-stepred-mediatedrecombinationforversatilehigh-efficiencymarkerlessdnamanipulationinescherichiacoli.biotechnol.tech.40,191-197.32.tischer,b.k.,kaufer,b.b.,sommer,m.,wussow,f.,arvin,a.,andosterrieder,n.aself-excisableinfectiousbacterialartificialchromosomecloneofvaricella-zostervirusallowsanalysisoftheessentialtegumentproteinencodedbyorf9.j.virol.81(23),2007,13200-13208.33.tischer,b.k,smith,g.a.,andosterrieder,n.in:jeffbraman(ed.),invitromutagenesisprotocols:thirdedition,methodsinmolecularbiology,vol.634,doi10.1007/978-1-60761-652-8_30,springerscience+businessmedia,llc2010,chapter30:enpassantmutagenesis:atwostepmarkerlessredrecombinationsystem.34.thompson,s.r.2012.tricksaniresusestoenslaveribosomes.trendsmicrobiol.nov;20(11):558-66.35.trapp,s.,voneinem,j.,hofmann,h.,kostler,j.,wild,j.,wagner,r.,beer,m.,osterrieder,n.,2005.potentialofequineherpesvirus1asavectorforimmunization.j.virol.79,5445-5454.36.trombettacm,perinid,mathers,tempertonn,montomolie.2014.overviewofserologicaltechniquesforinfluenzavaccineevaluation:past,presentandfuture.vaccines(basel)13;2(4):707-34.doi:10.3390/vaccines2040707.37.wellington,j.e.,allen,g.p.,gooley,a.a.,love,d.n.,packer,n.h.,yan,j.x.,whalley,j.m.1996.thehighlyo-glycosylatedglycoproteingp2ofequineherpesvirus1isencodedbygene71.jvirol.70(11):8195-8.38.wernikek,aebischera,roman-sosag,beerm,(2017).then-terminaldomainofschmallenbergvirusenvelopeproteingcishighlyimmunogenicandcanprovideprotectionfrominfection.scientificreports.2017feb13;7:42500.39.wernikek,eschbaumerm,breithaupta,hoffmannb,beerm(2012).schmallenbergviruschallengemodelsincattle:infectiousserumorculture-grownvirus?veterinaryresearch43,8440.wernikek,eschbaumerm,schirrmeierh,blohmu,breithaupta,hoffmannb,beerm,(2013a).oralexposure,reinfectionandcellularimmunitytoschmallenbergvirusincattle.veterinarymicrobiology165,155-15941.wernikek,nikolinvm,hechingers,hoffmannb,beerm(2013b).inactivatedschmallenbergvirusprototypevaccines.vaccine31,3558-3563序列表<110>勃林格殷格翰动物保健有限公司<120>新颖的启动子<130>p01-3202<160>37<170>patentinversion3.5<210>1<211>600<212>dna<213>equineherpesvirus4<400>1gcagactttggagcagcacaatttccggttgtggaccccatggaccttggtttggctggt60accgtggaaactaacgctccggaagttttggccagagcaaaatacaattcgaaggtagac120atatggagcgccggaatagttctgtttgaaatgctcgcatatccatcaactctatttgag180gacccgccgagtaccccacaagagtatgtaaaaagctgtcattctcaactactgagaata240atatcaaagctaaagataaaccctgaggagtttccacgggaaccagagtctaggctcgtg300cgcggatacatcgaatacgccagcctagagcgtaagccacatacgcgctatccttgcttc360cagcgcgtgaacctacacattgacggggaatttttgatccataaaatgctagcgttcaat420gctgcgatgcgcccatccgcagaagagttgttgtcctacccaatgtttatgaatctgtag480gatgactaacagatttggggtggagacggcgtgggcgatactgtataaagttgtactact540taccagcccagtcagtgtgctgtagtgccaccacctgtaaagctgtgataagctgcagtt600<210>2<211>600<212>dna<213>equineherpesvirus4<400>2agctgggggagtttgtactatagtgtattacatgcggcttgcaataactgcctggtttat60gtttcgcaacattcaagcagacatgctaccgctaaacactttgcaacaattttttattgg120gtgtttggcctttggtagaactgtcgcgtttttggtggtagcatatactaccttatttat180acgctccgagctgtttttcagcatgctagcacccaacgccgagcgagagtatataactcc240catcattgcccacaagcttatgccacttattagcgtccgctctgccgtttgcttagtcat300aatatctaccgccgtttacgcagcagacgctatctgcgacacaattggatttgcgatacc360gcgcatgtggatgtgtattttaatgagatcaacctccatgaagcgtaactagggggcctc420ccactgaggcactaccggcttagcagctgactaacacagtataaaacgtgagaagaaatc480agtctcatgcgccattagcgctaggctagttagcgtggaggaccggagcgctaccgccag540cagtttcatccgcctggttacgggtttgttaacacctaccggtgttttaccgctaccata600<210>3<211>430<212>dna<213>equineherpesvirus4<400>3tctatttgaggacccgccgagtaccccacaagagtatgtaaaaagctgtcattctcaact60actgagaataatatcaaagctaaagataaaccctgaggagtttccacgggaaccagagtc120taggctcgtgcgcggatacatcgaatacgccagcctagagcgtaagccacatacgcgcta180tccttgcttccagcgcgtgaacctacacattgacggggaatttttgatccataaaatgct240agcgttcaatgctgcgatgcgcccatccgcagaagagttgttgtcctacccaatgtttat300gaatctgtaggatgactaacagatttggggtggagacggcgtgggcgatactgtataaag360ttgtactacttaccagcccagtcagtgtgctgtagtgccaccacctgtaaagctgtgata420agctgcagtt430<210>4<211>449<212>dna<213>equineherpesvirus4<400>4ttggtggtagcatatactaccttatttatacgctccgagctgtttttcagcatgctagca60cccaacgccgagcgagagtatataactcccatcattgcccacaagcttatgccacttatt120agcgtccgctctgccgtttgcttagtcataatatctaccgccgtttacgcagcagacgct180atctgcgacacaattggatttgcgataccgcgcatgtggatgtgtattttaatgagatca240acctccatgaagcgtaactagggggcctcccactgaggcactaccggcttagcagctgac300taacacagtataaaacgtgagaagaaatcagtctcatgcgccattagcgctaggctagtt360agcgtggaggaccggagcgctaccgccagcagtttcatccgcctggttacgggtttgtta420acacctaccggtgttttaccgctaccata449<210>5<211>20<212>dna<213>artificialsequence<220><223>primerno1130specificfororf72<400>5tgtctaccttcaagcttatg20<210>6<211>17<212>dna<213>artificialsequence<220><223>primerno1131specificfororf72<400>6ctagcgcagtcgcgttg17<210>7<211>18<212>dna<213>artificialsequence<220><223>primerno.1079specificformcherry<400>7gcgaggaggataacatgg18<210>8<211>18<212>dna<213>artificialsequence<220><223>primerno.1080specificformcherry<400>8acccttggtcaccttcag18<210>9<211>22<212>dna<213>artificialsequence<220><223>artificialsequencenucleicacidpcrprimer1017fortheorf70insertionregion<400>9aggctcgtgcgcggatacatcg22<210>10<211>22<212>dna<213>artificialsequence<220><223>artificialsequencenucleicacidpcrprimer1018fortheorf70insertionregion<400>10ttcggggctgttagactcctcc22<210>11<211>22<212>dna<213>artificialsequence<220><223>artificialsequencenucleicacidpcrprimer1007fortheorf1/3insertionregion<400>11ccaactcgccgccatgagaccc22<210>12<211>22<212>dna<213>artificialsequence<220><223>artificialsequencenucleicacidpcrprimer1008fortheorf1/3insertionregion<400>12agcgcgccccgtacccagtggg22<210>13<211>417<212>dna<213>equineherpesvirus1<400>13ctccgagtaccccagaggagtatgtgaaaagctgccactcgcaactactgaagataattt60caacgctcaagataaatccggaggagtttcctcgagaccccgggtcgaggctcgtgcgcg120gatacatcgagtattctagactcgagcgcaagccctacacgcgctacccctgctttcaac180gcgtcaacctgcacattgacggggagtttctggttcacaagatgctagcgttcaatgccg240cgatgcgcccatcggccgaggagctgctgtcatacccaatgtttgctcaactttaggatg300actaacctgtttctgggaggagacagcgtgggcgacggtgtataaagttggtctgctttc360aagccctgccactgcgctacagtgccaccaactgtaaagcggtagtaagctgcagtg417<210>14<211>431<212>dna<213>equineherpesvirus1<400>14gaccctgttggtgggtgcggttggactcagaatcttggcgcaggcatggaagtttgtcgg60tgacgaaacatacgacaccatccgcgcagaagcaaagaatttagagacccacgtaccctc120aagtgctgcagagtcgtctctagaaaaccaatcgacacaggaggagtctaacagccccga180agttgcccacctgcgaagcgtcaacagcgatgacagtacacacacggggggtgcgtcgaa240cggcatccaggactgtgacagtcagctcaaaactgtgtatgcctgcttggctctaattgg300actcggcacatgtgccatgatagggttgatagtttacatttgtgtattaaggtcaaaact360gtcctctcggaatttttcgcgcgcgcaaaatgtaaaacatagaaattaccagcgacttga420gtacgttgctt431<210>15<211>417<212>dna<213>equineherpesvirus1<400>15ctccgagtaccccagaggagtatgtgaaaagctgccactcgcaactactgaagataattt60caacgctcaagataaatccggaggagtttcctcgagaccccgggtcgaggctcgtgcgcg120gatacatcgagtattctagactcgagcgcaagccctacacgcgctacccctgctttcaac180gcgtcaacctgcacattgacggggagtttctggttcacaagatgctagcgttcaatgccg240cgatgcgcccatcggccgaggagctgctgtcatacccaatgtttgcacaactttaggatg300actaacctgtttctgggaggagacagcgtgggcgacggtgtataaagttggtctgctttc360aagccctgccactgcgctacagtgccaccaactgtaaagcggtagtaagctgcagtg417<210>16<211>431<212>dna<213>equineherpesvirus1<400>16gaccctgttggtgggtgcggttggactcagaatcttggcgcaggcatggaagtttgtcgg60tgacgaaacatacgacaccatccgcgcagaagcaaagaatttagagacccacgtaccctc120aagtgctgcagagtcgtctctagaaaaccaatcgacacaggaggagtctaacagccccga180agttgcccacctgcgaagcgtcaacagcgatgacagtacacacacggggggtgcgtcgaa240cggcatccaggactgtgacagtcagctcaaaactgtgtatgcctgcttggctctaattgg300actcggcacatgtgccatgatagggttgatagtttacatttgtgtattaaggtcaaaact360gtcctctcggaatttttcgcgcgcgcaaaatgtaaaacatagaaattaccagcgacttga420gtacgttgctt431<210>17<211>283<212>dna<213>equineherpesvirus1<400>17tctagactcgagcgcaagccctacacgcgctacccctgctttcaacgcgtcaacctgcac60attgacggggagtttctggttcacaagatgctagcgttcaatgccgcgatgcgcccatcg120gccgaggagctgctgtcatacccaatgtttgctcaactttaggatgactaacctgtttct180gggaggagacagcgtgggcgacggtgtataaagttggtctgctttcaagccctgccactg240cgctacagtgccaccaactgtaaagcggtagtaagctgcagtg283<210>18<211>144<212>dna<213>equineherpesvirus1<400>18gaccctgttggtgggtgcggttggactcagaatcttggcgcaggcatggaagtttgtcgg60tgacgaaacatacgacaccatccgcgcagaagcaaagaatttagagacccacgtaccctc120aagtgctgcagagtcgtctctaga144<210>19<211>801<212>dna<213>equineherpesvirus1<400>19atgttgactgtcttagcagccctgagtctgctcagcttgcttacgagcgcaaccggacgg60ctcgccccagatgaactctgttatgccgaaccccgcagaactggcagcccaccaaacacc120cagcccgaacgcccacccgtaatatttgagcccccaacaattgcgattaaagctgaatcc180aagggttgtgagctaattttattagatccacccatagatgtaagctatcgcagagaagat240aaggtgaatgcgtccattgcttggttttttgactttggcgcttgccggatgcccatcgca300tacagagagtattacggttgtattggcaatgctgttccctccccagagacttgtgatgcg360tactcatttacccttattaggaccgagggtatcgtggagtttaccatcgtaaacatgagc420ctcctgtttcagcctggaatatacgatagtggcaattttatctacagcgttctcctggac480taccacatatttacaggacgtgtaacgttggaagtggaaaaggacacaaactatccctgt540ggcatgattcatggactcactgcttacggaaacatcaacgtagatgaaaccatggacaac600gccagcccacacccgcgtgccgtggggtgctttcccgagcccatcgacaacgaagcgtgg660gcaaacgttacatttactgaattggggataccagacccaaactcatttctcgatgacgag720ggtgattacccgaatatatcagactgtcactcgtgggagtcatacacctacccaaatacg780ctgaggcaggccacaggaccc801<210>20<211>801<212>dna<213>equineherpesvirus1<400>20atgttgactgtcttagcagctctgagtctgctcagcttgcttacgagcgcaaccggacgg60ctcgccccagatgaactctgttatgccgaaccccgcagaactggcagcccaccaaacacc120cagcccgaacgcccacccgtaatatttgagcccccaacaattgcgattaaagctgaatcc180aagggttgtgagctaattttattagatccacccatagatgtaagctatcgcagagaagat240aaggtgaatgcgtccattgcttggttttttgactttggcgcttgccggatgcccatcgca300tacagagagtattacggttgtattggcaatgctgttccctccccagagacttgtgatgcg360tactcatttacccttattaggaccgagggtatcgtggagtttaccatcgtaaacatgagc420ctcctgtttcagcctggaatatacgatagtggcaattttatctacagcgttctcctggac480taccacatatttacaggacgtgtaacgttggaagtggaaaaggacacaaactatccctgt540ggcatgattcatggactcactgcttacggaaacatcaacgtagatgaaaccatggacaac600gccagcccacacccgcgtgccgtggggtgctttcccgagcccatcgacaacgaagcgtgg660gcaaacgttacatttactgaattggggataccagacccaaactcatttctcgatgacgag720ggtgattacccgaatatatcagactgtcactcgtgggagtcatacacctacccaaatacg780ctgaggcaggccacaggaccc801<210>21<211>4435<212>dna<213>artificialsequence<220><223>nucleotidesequenceoftransferplasmidpu-mc70-bgh<400>21tcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtca60cagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtg120ttggcgggtgtcggggctggcttaactatgcggcatcagagcagattgtactgagagtgc180accatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgcc240attcgccattcaggctgcgcaactgttgggaagggcgatcggtgcgggcctcttcgctat300tacgccagctggcgaaagggggatgtgctgcaaggcgattaagttgggtaacgccagggt360tttcccagtcacgacgttgtaaaacgacggccagtgaattcctccgagtaccccagagga420gtatgtgaaaagctgccactcgcaactactgaagataatttcaacgctcaagataaatcc480ggaggagtttcctcgagaccccgggtcgaggctcgtgcgcggatacatcgagtattctag540actcgagcgcaagccctacacgcgctacccctgctttcaacgcgtcaacctgcacattga600cggggagtttctggttcacaagatgctagcgttcaatgccgcgatgcgcccatcggccga660ggagctgctgtcatacccaatgtttgctcaactttaggatgactaacctgtttctgggag720gagacagcgtgggcgacggtgtataaagttggtctgctttcaagccctgccactgcgcta780cagtgccaccaactgtaaagcggtagtaagctgcagtggtcgacatggtgagcaagggcg840aggaggataacatggccatcatcaaggagttcatgcgcttcaaggtgcacatggagggct900ccgtgaacggccacgagttcgagatcgagggcgagggcgagggccgcccctacgagggca960cccagaccgccaagctgaaggtgaccaagggtggccccctgcccttcgcctgggacatcc1020tgtcccctcagttcatgtacggctccaaggcctacgtgaagcaccccgccgacatccccg1080actacttgaagctgtccttccccgagggcttcaagtgggagcgcgtgatgaacttcgagg1140acggcggcgtggtgaccgtgacccaggactcctccctgcaggacggcgagttcatctaca1200aggtgaagctgcgcggcaccaacttcccctccgacggccccgtaatgcagaagaagacca1260tgggctgggaggcctcctccgagcggatgtaccccgaggacggcgccctgaagggcgaga1320tcaagcagaggctgaagctgaaggacggcggccactacgacgctgaggtcaagaccacct1380acaaggccaagaagcccgtgcagctgcccggcgcctacaacgtcaacatcaagttggaca1440tcacctcccacaacgaggactacaccatcgtggaacagtacgaacgcgccgagggccgcc1500actccaccggcggcatggacgagctgtacaagtaactgtgccttctagttgccagccatc1560tgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcct1620ttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggg1680gggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctgg1740ggatgcggtgggctctatggatccgaccctgttggtgggtgcggttggactcagaatctt1800ggcgcaggcatggaagtttgtcggtgacgaaacatacgacaccatccgcgcagaagcaaa1860gaatttagagacccacgtaccctcaagtgctgcagagtcgtctctagaaaaccaatcgac1920acaggaggagtctaacagccccgaagttgcccacctgcgaagcgtcaacagcgatgacag1980tacacacacggggggtgcgtcgaacggcatccaggactgtgacagtcagctcaaaactgt2040gtatgcctgcttggctctaattggactcggcacatgtgccatgatagggttgatagttta2100catttgtgtattaaggtcaaaactgtcctctcggaatttttcgcgcgcgcaaaatgtaaa2160acatagaaattaccagcgacttgagtacgttgcttaagcttggcgtaatcatggtcatag2220ctgtttcctgtgtgaaattgttatccgctcacaattccacacaacatacgagccggaagc2280ataaagtgtaaagcctggggtgcctaatgagtgagctaactcacattaattgcgttgcgc2340tcactgcccgctttccagtcgggaaacctgtcgtgccagctgcattaatgaatcggccaa2400cgcgcggggagaggcggtttgcgtattgggcgctcttccgcttcctcgctcactgactcg2460ctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacgg2520ttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaag2580gccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgac2640gagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaaga2700taccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgctt2760accggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgc2820tgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccc2880cccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggta2940agacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtat3000gtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggaca3060gtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctct3120tgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagatt3180acgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgct3240cagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttc3300acctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaa3360acttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtcta3420tttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggc3480ttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagat3540ttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaacttta3600tccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagtt3660aatagtttgcgcaacgttgttgccattgctacaggcatcgtggtgtcacgctcgtcgttt3720ggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatg3780ttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggcc3840gcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatcc3900gtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatg3960cggcgaccgagttgctcttgcccggcgtcaatacgggataataccgcgccacatagcaga4020actttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatctta4080ccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatct4140tttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaag4200ggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattga4260agcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaat4320aaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaacc4380attattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtc4435<210>22<211>5191<212>dna<213>artificialsequence<220><223>nucleotidesequenceoftransfervectorpu70-p455-71k71<400>22caataaacgcggtatgtctaccttcaagcctatgatgaacggatgtttggtgtttgcggc60tattataacgctcttgagttttatgctatctctgggaacatgcgaaaattacaggcgtgt120ggttcgggatcctagggataacagggtaatcgatttattcaacaaagccacgttgtgtct180caaaatctctgatgttacattgcacaagataaaaatatatcatcatgaacaataaaactg240tctgcttacataaacagtaatacaaggggtgttatgagccatattcaacgggaaacgtct300tgctcgaggccgcgattaaattccaacatggatgctgatttatatgggtataaatgggct360cgcgataatgtcgggcaatcaggtgcgacaatctatcgattgtatgggaagcccgatgcg420ccagagttgtttctgaaacatggcaaaggtagcgttgccaatgatgttacagatgagatg480gtcagactaaactggctgacggaatttatgcctcttccgaccatcaagcattttatccgt540actcctgatgatgcatggttactcaccactgcgatccccgggaaaacagcattccaggta600ttagaagaatatcctgattcaggtgaaaatattgttgatgcgctggcagtgttcctgcgc660cggttgcattcgattcctgtttgtaattgtccttttaacagcgatcgcgtatttcgtctc720gctcaggcgcaatcacgaatgaataacggtttggttgatgcgagtgattttgatgacgag780cgtaatggctggcctgttgaacaagtctggaaagaaatgcataagcttttgccattctca840ccggattcagtcgtcactcatggtgatttctcacttgataaccttatttttgacgagggg900aaattaataggttgtattgatgttggacgagtcggaatcgcagaccgataccaggatctt960gccatcctatggaactgcctcggtgagttttctccttcattacagaaacggctttttcaa1020aaatatggtattgataatcctgatatgaataaattgcagtttcatttgatgctcgatgag1080tttttctaaaataaacgcggtatgtctaccttcaagcctatgatgaacggatgtttggtg1140tttgcggctattataacgctcttgagttttatgctatctctgggaacatgcgaaaattac1200aggcgtgtggttcgggatccgaccctgttggtgggtgcggttggactcagaatcttggcg1260caggcatggaagtttgtcggtgacgaaacatacgacaccatccgcgcagaagcaaagaat1320ttagagacccacgtaccctcaagtgctgcagagtcgtctctagaaaaccaatcgacacag1380gaggagtctaacagccccgaagttgcccacctgcgaagcgtcaacagcgatgacagtaca1440cacacggggggtgcgtcgaacggcatccaggactgtgacagtcagctcaaaactgtgtat1500gcctgcttggctctaattggactcggcacatgtgccatgatagggttgatagtttacatt1560tgtgtattaaggtcaaaactgtcctctcggaatttttcgcgcgcgcaaaatgtaaaacat1620agaaattaccagcgacttgagtacgttgcttaagcttggcgtaatcatggtcatagctgt1680ttcctgtgtgaaattgttatccgctcacaattccacacaacatacgagccggaagcataa1740agtgtaaagcctggggtgcctaatgagtgagctaactcacattaattgcgttgcgctcac1800tgcccgctttccagtcgggaaacctgtcgtgccagctgcattaatgaatcggccaacgcg1860cggggagaggcggtttgcgtattgggcgctcttccgcttcctcgctcactgactcgctgc1920gctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttat1980ccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggcca2040ggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagc2100atcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagatacc2160aggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccg2220gatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgta2280ggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccg2340ttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagac2400acgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtag2460gcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtat2520ttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgat2580ccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgc2640gcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagt2700ggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacct2760agatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaactt2820ggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttc2880gttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttac2940catctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttat3000cagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccg3060cctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaata3120gtttgcgcaacgttgttgccattgctacaggcatcgtggtgtcacgctcgtcgtttggta3180tggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgt3240gcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcag3300tgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaa3360gatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggc3420gaccgagttgctcttgcccggcgtcaatacgggataataccgcgccacatagcagaactt3480taaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgc3540tgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatctttta3600ctttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaa3660taagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagca3720tttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaac3780aaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccatta3840ttatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtctcgcgcgtt3900tcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtc3960tgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggt4020gtcggggctggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgc4080ggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgccattcgccat4140tcaggctgcgcaactgttgggaagggcgatcggtgcgggcctcttcgctattacgccagc4200tggcgaaagggggatgtgctgcaaggcgattaagttgggtaacgccagggttttcccagt4260cacgacgttgtaaaacgacggccagtgaattcctccgagtaccccagaggagtatgtgaa4320aagctgccactcgcaactactgaagataatttcaacgctcaagataaatccggaggagtt4380tcctcgagaccccgggtcgaggctcgtgcgcggatacatcgagtattctagactcgagcg4440caagccctacacgcgctacccctgctttcaacgcgtcaacctgcacattgacggggagtt4500tctggttcacaagatgctagcgttcaatgccgcgatgcgcccatcggccgaggagctgct4560gtcatacccaatgtttgctcaactttaggatgactaacctgtttctgggaggagacagcg4620tgggcgacggtgtataaagttggtctgctttcaagccctgccactgcgctacagtgccac4680caactgtaaagcggtagtaagctgcagtggtcgactggtggtagcatatactaccttatt4740tatacgctccgagctgtttttcagcatgctagcacccaacgccgagcgagagtatataac4800tcccatcattgcccacaagcttatgccacttattagcgtccgctctgccgtttgcttagt4860cataatatctaccgccgtttacgcagcagacgctatctgcgacacaattggatttgcgat4920accgcgcatgtggatgtgtattttaatgagatcaacctccatgaagcgtaactagggggc4980ctcccactgaggcactaccggcttagcagctgactaacacagtataaaacgtgagaagaa5040atcagtctcatgcgccattagcgctaggctagttagcgtggaggaccggagcgctaccgc5100cagcagtttcatccgcctggttacgggtttgttaacacctaccggtgttttaccgctacc5160ataggatccgatccatgggcggccgcggtac5191<210>23<211>6892<212>dna<213>artificialsequence<220><223>nucleotidesequenceoftransferplasmidpu70-p455-h3-71k71<400>23caataaacgcggtatgtctaccttcaagcctatgatgaacggatgtttggtgtttgcggc60tattataacgctcttgagttttatgctatctctgggaacatgcgaaaattacaggcgtgt120ggttcgggatcctagggataacagggtaatcgatttattcaacaaagccacgttgtgtct180caaaatctctgatgttacattgcacaagataaaaatatatcatcatgaacaataaaactg240tctgcttacataaacagtaatacaaggggtgttatgagccatattcaacgggaaacgtct300tgctcgaggccgcgattaaattccaacatggatgctgatttatatgggtataaatgggct360cgcgataatgtcgggcaatcaggtgcgacaatctatcgattgtatgggaagcccgatgcg420ccagagttgtttctgaaacatggcaaaggtagcgttgccaatgatgttacagatgagatg480gtcagactaaactggctgacggaatttatgcctcttccgaccatcaagcattttatccgt540actcctgatgatgcatggttactcaccactgcgatccccgggaaaacagcattccaggta600ttagaagaatatcctgattcaggtgaaaatattgttgatgcgctggcagtgttcctgcgc660cggttgcattcgattcctgtttgtaattgtccttttaacagcgatcgcgtatttcgtctc720gctcaggcgcaatcacgaatgaataacggtttggttgatgcgagtgattttgatgacgag780cgtaatggctggcctgttgaacaagtctggaaagaaatgcataagcttttgccattctca840ccggattcagtcgtcactcatggtgatttctcacttgataaccttatttttgacgagggg900aaattaataggttgtattgatgttggacgagtcggaatcgcagaccgataccaggatctt960gccatcctatggaactgcctcggtgagttttctccttcattacagaaacggctttttcaa1020aaatatggtattgataatcctgatatgaataaattgcagtttcatttgatgctcgatgag1080tttttctaaaataaacgcggtatgtctaccttcaagcctatgatgaacggatgtttggtg1140tttgcggctattataacgctcttgagttttatgctatctctgggaacatgcgaaaattac1200aggcgtgtggttcgggatccgaccctgttggtgggtgcggttggactcagaatcttggcg1260caggcatggaagtttgtcggtgacgaaacatacgacaccatccgcgcagaagcaaagaat1320ttagagacccacgtaccctcaagtgctgcagagtcgtctctagaaaaccaatcgacacag1380gaggagtctaacagccccgaagttgcccacctgcgaagcgtcaacagcgatgacagtaca1440cacacggggggtgcgtcgaacggcatccaggactgtgacagtcagctcaaaactgtgtat1500gcctgcttggctctaattggactcggcacatgtgccatgatagggttgatagtttacatt1560tgtgtattaaggtcaaaactgtcctctcggaatttttcgcgcgcgcaaaatgtaaaacat1620agaaattaccagcgacttgagtacgttgcttaagcttggcgtaatcatggtcatagctgt1680ttcctgtgtgaaattgttatccgctcacaattccacacaacatacgagccggaagcataa1740agtgtaaagcctggggtgcctaatgagtgagctaactcacattaattgcgttgcgctcac1800tgcccgctttccagtcgggaaacctgtcgtgccagctgcattaatgaatcggccaacgcg1860cggggagaggcggtttgcgtattgggcgctcttccgcttcctcgctcactgactcgctgc1920gctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttat1980ccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggcca2040ggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagc2100atcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagatacc2160aggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccg2220gatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgta2280ggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccg2340ttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagac2400acgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtag2460gcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtat2520ttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgat2580ccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgc2640gcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagt2700ggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacct2760agatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaactt2820ggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttc2880gttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttac2940catctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttat3000cagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccg3060cctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaata3120gtttgcgcaacgttgttgccattgctacaggcatcgtggtgtcacgctcgtcgtttggta3180tggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgt3240gcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcag3300tgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaa3360gatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggc3420gaccgagttgctcttgcccggcgtcaatacgggataataccgcgccacatagcagaactt3480taaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgc3540tgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatctttta3600ctttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaa3660taagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagca3720tttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaac3780aaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccatta3840ttatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtctcgcgcgtt3900tcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtc3960tgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggt4020gtcggggctggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgc4080ggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgccattcgccat4140tcaggctgcgcaactgttgggaagggcgatcggtgcgggcctcttcgctattacgccagc4200tggcgaaagggggatgtgctgcaaggcgattaagttgggtaacgccagggttttcccagt4260cacgacgttgtaaaacgacggccagtgaattcctccgagtaccccagaggagtatgtgaa4320aagctgccactcgcaactactgaagataatttcaacgctcaagataaatccggaggagtt4380tcctcgagaccccgggtcgaggctcgtgcgcggatacatcgagtattctagactcgagcg4440caagccctacacgcgctacccctgctttcaacgcgtcaacctgcacattgacggggagtt4500tctggttcacaagatgctagcgttcaatgccgcgatgcgcccatcggccgaggagctgct4560gtcatacccaatgtttgctcaactttaggatgactaacctgtttctgggaggagacagcg4620tgggcgacggtgtataaagttggtctgctttcaagccctgccactgcgctacagtgccac4680caactgtaaagcggtagtaagctgcagtggtcgactggtggtagcatatactaccttatt4740tatacgctccgagctgtttttcagcatgctagcacccaacgccgagcgagagtatataac4800tcccatcattgcccacaagcttatgccacttattagcgtccgctctgccgtttgcttagt4860cataatatctaccgccgtttacgcagcagacgctatctgcgacacaattggatttgcgat4920accgcgcatgtggatgtgtattttaatgagatcaacctccatgaagcgtaactagggggc4980ctcccactgaggcactaccggcttagcagctgactaacacagtataaaacgtgagaagaa5040atcagtctcatgcgccattagcgctaggctagttagcgtggaggaccggagcgctaccgc5100cagcagtttcatccgcctggttacgggtttgttaacacctaccggtgttttaccgctacc5160ataggatccgatccatgggcggccgcatgaagaccgtgatcgccctgagttacatcttct5220gcctggtgtttgggcaggacctccctggtaaaggcaacaacacggccacgctgtgccttg5280ggcaccacgccgtgccgaacggcacccttgtgaaaactattaccgacgatcagatcgagg5340tgaccaacgccaccgaactggttcagaattttagcatgggcaaaatttgcaataacccgc5400accgcattctggacggggccaactgcacgctgatcgattcattgctgggtgatccccact5460gcgatggctttcaaaacgaaaagtgggacttgttcatcgaacgcagcaaggcattcagca5520actgctacccatacgacgtgcccgaatacaccagcctgcgaagcctgatcgcgagctctg5580ggaccctggagttcaccaatgagaacttcaattggaccggagtgacccaaaacggtggct5640ccagcgcctgtaaaaggggacccaataacagcttctttagcaagttgaattggctttaca5700agagcggcaatacttacccgatgttgaatgtgaccatgcccaacagtgacgactttgata5760aactgtacatatggggcgtgcaccatcccagcacggaccgcgaacagataaacctgtacg5820tgcaggccagcgggaagataatcgtgagcaccaagcgcagccagcagaccatcattccca5880acattggcagccgaccgtgggtgcgcggtctgagctcccgcatcagcatatactggacca5940ttgtcaagccgggagacatcctgatcatcaactctaatggcaatcttatcgccccacgcg6000gctacttcaagatgcagaccggcaaaagcagtgtgatgaggagcgacgcccccatcgaca6060cctgcaatagcgaatgcatcacccccaatggcagcatccccaacgacaagcctttccaga6120acgtgaataagatcacctacggcgcgtgccccaagtacatcaagcagaacaccctgaagc6180tggccaccggcatgcgcaacatccccgagcgacagacacggggcatttttggcgcaatcg6240cagggttcattgagaatggctgggagggaatggttaacggctggtacggcttccgccatc6300agaactctgaaggaatcggccaagctgcggatctgaagtccacgcaagcagccatcaacc6360agatcaacggcaagcttaaccgcgtgattgaaaagacgaacgagaaattccaccaaatag6420agaaagaattcagcgaggtggagggccgcatccaagacctcgagcgctacgtggaggaca6480ccaagatcgacctgtggagctacaatgccgagctcctggtcgccttggaaaaccaacaca6540ccattgacctgaccgacagcgagatgaataaactcttcgagaagacccggaagcaactcc6600gagagaacgccgaagacatgggtaatgggtgttttaagatctaccacaagtgcgacaata6660gctgcatggagagcatccgaaacggaacctacgaccacaacgagtaccgcgatgaggcag6720ttaataaccgcttccaaatcaaaagcgtggaactgaagagtggctataaggactggatac6780tgtggatcagctttgccataagctgcttcctgctgtgcgccgtttggttgggtttcatca6840tgtgggcctgtcaaaagggcaatattcgctgtaacatctgcatttgaggtac6892<210>24<211>4977<212>dna<213>artificialsequence<220><223>nucleotidesequenceoftransfervectorpu-1-3-p430-bghkbgh<400>24cctgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgacc60ctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgt120ctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggat180tgggaagacaatagcaggcatgctggggatgcggtgggctctatggatcctagggataac240agggtaatcgatttattcaacaaagccacgttgtgtctcaaaatctctgatgttacattg300cacaagataaaaatatatcatcatgaacaataaaactgtctgcttacataaacagtaata360caaggggtgttatgagccatattcaacgggaaacgtcttgctcgaggccgcgattaaatt420ccaacatggatgctgatttatatgggtataaatgggctcgcgataatgtcgggcaatcag480gtgcgacaatctatcgattgtatgggaagcccgatgcgccagagttgtttctgaaacatg540gcaaaggtagcgttgccaatgatgttacagatgagatggtcagactaaactggctgacgg600aatttatgcctcttccgaccatcaagcattttatccgtactcctgatgatgcatggttac660tcaccactgcgatccccgggaaaacagcattccaggtattagaagaatatcctgattcag720gtgaaaatattgttgatgcgctggcagtgttcctgcgccggttgcattcgattcctgttt780gtaattgtccttttaacagcgatcgcgtatttcgtctcgctcaggcgcaatcacgaatga840ataacggtttggttgatgcgagtgattttgatgacgagcgtaatggctggcctgttgaac900aagtctggaaagaaatgcataagcttttgccattctcaccggattcagtcgtcactcatg960gtgatttctcacttgataaccttatttttgacgaggggaaattaataggttgtattgatg1020ttggacgagtcggaatcgcagaccgataccaggatcttgccatcctatggaactgcctcg1080gtgagttttctccttcattacagaaacggctttttcaaaaatatggtattgataatcctg1140atatgaataaattgcagtttcatttgatgctcgatgagtttttctaaccatggctgtgcc1200ttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaagg1260tgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtag1320gtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaaga1380caatagcaggcatgctggggatgcggtgggctctatggatccgaccctccccggggctaa1440aaagctgcgtcttcacgcccgaggcgcttattgcccactgggtacggggcgcgcttttat1500atgtgtaacgtcccaccggtgtgacgcacgtactacggttgttctaaatagctgtccccg1560tgattgcctcggctgcacacatcgcctaggtttccgccgtgcctggtgtcgagggcccac1620ccctgtaaccaacatcgatgggggcctgctgctccttcgctaccttaggaccgttatagt1680tacgtcaagcttggcgtaatcatggtcatagctgtttcctgtgtgaaattgttatccgct1740cacaattccacacaacatacgagccggaagcataaagtgtaaagcctggggtgcctaatg1800agtgagctaactcacattaattgcgttgcgctcactgcccgctttccagtcgggaaacct1860gtcgtgccagctgcattaatgaatcggccaacgcgcggggagaggcggtttgcgtattgg1920gcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagc1980ggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcagg2040aaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgct2100ggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtca2160gaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccct2220cgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttc2280gggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgt2340tcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatc2400cggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagc2460cactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtg2520gtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagcc2580agttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtag2640cggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaaga2700tcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggat2760tttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaag2820ttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaat2880cagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccc2940cgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgat3000accgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaag3060ggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttg3120ccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgc3180tacaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttccca3240acgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcgg3300tcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagc3360actgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagta3420ctcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtc3480aatacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacg3540ttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacc3600cactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagc3660aaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaat3720actcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgag3780cggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttcc3840ccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaa3900taggcgtatcacgaggccctttcgtctcgcgcgtttcggtgatgacggtgaaaacctctg3960acacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagaca4020agcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggctggcttaactatgcggc4080atcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgt4140aaggagaaaataccgcatcaggcgccattcgccattcaggctgcgcaactgttgggaagg4200gcgatcggtgcgggcctcttcgctattacgccagctggcgaaagggggatgtgctgcaag4260gcgattaagttgggtaacgccagggttttcccagtcacgacgttgtaaaacgacggccag4320tgaattcgacgtaactataacggtcctaaggtagcgaatttttccattgggcccctccct4380tttggctctgggtatttagcttccctcccacttctcattccactttctccacctgcacct4440tttccatctcctctccaactcgccgccatgagacccgagggagtttcgcggggccgcgcc4500tcctctgtctccatctccaactagtgtcgacctctatttgaggacccgccgagtacccca4560caagagtatgtaaaaagctgtcattctcaactactgagaataatatcaaagctaaagata4620aaccctgaggagtttccacgggaaccagagtctaggctcgtgcgcggatacatcgaatac4680gccagcctagagcgtaagccacatacgcgctatccttgcttccagcgcgtgaacctacac4740attgacggggaatttttgatccataaaatgctagcgttcaatgctgcgatgcgcccatcc4800gcagaagagttgttgtcctacccaatgtttatgaatctgtaggatgactaacagatttgg4860ggtggagacggcgtgggcgatactgtataaagttgtactacttaccagcccagtcagtgt4920gctgtagtgccaccacctgtaaagctgtgataagctgcagttgcggccgccgggtac4977<210>25<211>6678<212>dna<213>artificialsequence<220><223>nucleotidesequenceoftransferplasmidpu1-3-p430-h1av-bghkbgh<400>25cctgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgacc60ctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgt120ctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggat180tgggaagacaatagcaggcatgctggggatgcggtgggctctatggatcctagggataac240agggtaatcgatttattcaacaaagccacgttgtgtctcaaaatctctgatgttacattg300cacaagataaaaatatatcatcatgaacaataaaactgtctgcttacataaacagtaata360caaggggtgttatgagccatattcaacgggaaacgtcttgctcgaggccgcgattaaatt420ccaacatggatgctgatttatatgggtataaatgggctcgcgataatgtcgggcaatcag480gtgcgacaatctatcgattgtatgggaagcccgatgcgccagagttgtttctgaaacatg540gcaaaggtagcgttgccaatgatgttacagatgagatggtcagactaaactggctgacgg600aatttatgcctcttccgaccatcaagcattttatccgtactcctgatgatgcatggttac660tcaccactgcgatccccgggaaaacagcattccaggtattagaagaatatcctgattcag720gtgaaaatattgttgatgcgctggcagtgttcctgcgccggttgcattcgattcctgttt780gtaattgtccttttaacagcgatcgcgtatttcgtctcgctcaggcgcaatcacgaatga840ataacggtttggttgatgcgagtgattttgatgacgagcgtaatggctggcctgttgaac900aagtctggaaagaaatgcataagcttttgccattctcaccggattcagtcgtcactcatg960gtgatttctcacttgataaccttatttttgacgaggggaaattaataggttgtattgatg1020ttggacgagtcggaatcgcagaccgataccaggatcttgccatcctatggaactgcctcg1080gtgagttttctccttcattacagaaacggctttttcaaaaatatggtattgataatcctg1140atatgaataaattgcagtttcatttgatgctcgatgagtttttctaaccatggctgtgcc1200ttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaagg1260tgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtag1320gtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaaga1380caatagcaggcatgctggggatgcggtgggctctatggatccgaccctccccggggctaa1440aaagctgcgtcttcacgcccgaggcgcttattgcccactgggtacggggcgcgcttttat1500atgtgtaacgtcccaccggtgtgacgcacgtactacggttgttctaaatagctgtccccg1560tgattgcctcggctgcacacatcgcctaggtttccgccgtgcctggtgtcgagggcccac1620ccctgtaaccaacatcgatgggggcctgctgctccttcgctaccttaggaccgttatagt1680tacgtcaagcttggcgtaatcatggtcatagctgtttcctgtgtgaaattgttatccgct1740cacaattccacacaacatacgagccggaagcataaagtgtaaagcctggggtgcctaatg1800agtgagctaactcacattaattgcgttgcgctcactgcccgctttccagtcgggaaacct1860gtcgtgccagctgcattaatgaatcggccaacgcgcggggagaggcggtttgcgtattgg1920gcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagc1980ggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcagg2040aaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgct2100ggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtca2160gaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccct2220cgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttc2280gggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgt2340tcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatc2400cggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagc2460cactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtg2520gtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagcc2580agttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtag2640cggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaaga2700tcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggat2760tttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaag2820ttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaat2880cagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccc2940cgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgat3000accgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaag3060ggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttg3120ccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgc3180tacaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttccca3240acgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcgg3300tcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagc3360actgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagta3420ctcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtc3480aatacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacg3540ttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacc3600cactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagc3660aaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaat3720actcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgag3780cggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttcc3840ccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaa3900taggcgtatcacgaggccctttcgtctcgcgcgtttcggtgatgacggtgaaaacctctg3960acacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagaca4020agcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggctggcttaactatgcggc4080atcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgt4140aaggagaaaataccgcatcaggcgccattcgccattcaggctgcgcaactgttgggaagg4200gcgatcggtgcgggcctcttcgctattacgccagctggcgaaagggggatgtgctgcaag4260gcgattaagttgggtaacgccagggttttcccagtcacgacgttgtaaaacgacggccag4320tgaattcgacgtaactataacggtcctaaggtagcgaatttttccattgggcccctccct4380tttggctctgggtatttagcttccctcccacttctcattccactttctccacctgcacct4440tttccatctcctctccaactcgccgccatgagacccgagggagtttcgcggggccgcgcc4500tcctctgtctccatctccaactagtgtcgacctctatttgaggacccgccgagtacccca4560caagagtatgtaaaaagctgtcattctcaactactgagaataatatcaaagctaaagata4620aaccctgaggagtttccacgggaaccagagtctaggctcgtgcgcggatacatcgaatac4680gccagcctagagcgtaagccacatacgcgctatccttgcttccagcgcgtgaacctacac4740attgacggggaatttttgatccataaaatgctagcgttcaatgctgcgatgcgcccatcc4800gcagaagagttgttgtcctacccaatgtttatgaatctgtaggatgactaacagatttgg4860ggtggagacggcgtgggcgatactgtataaagttgtactacttaccagcccagtcagtgt4920gctgtagtgccaccacctgtaaagctgtgataagctgcagttgcggccgccgatggaggc4980aaaattgttcgtgctgttctgcgccttcactgctctgaaggcagacaccatctgcgtggg5040ttaccacgccaataattccaccgacacggtggataccatcctggagaagaacgtgaccgt5100gactcattccgtgaacctcttggagaactcacacaatggtaaattgtgcagccttaacgg5160caaagccccgctgcaattggggaattgtaacgtggccggatggatactggggaaccccga5220gtgcgaccttctcctgaccgccaacagttggtcctacatcattgagacgagcaacagcaa5280gaatggcgcctgctatcctggggagttcgctgactacgaggagctgcgcgagcagttgtc5340tacagtcagcagcttcgaaagattcgagatcttcccaaaggccactagctggcccaacca5400cgatactaccaagggcactacagtgagttgcagccacagcggtgccaatagcttctaccg5460caacctgctgtggatcgtgaagaagggtaacagctaccccaagctgagcaaatcttacac5520aaacaacaaaggcaaagaggtgttggttatctggggcgtgcatcatcccccaaccgactc5580cgatcagcaaaccctgtaccagaacaaccacacctacgtgagcgtcggtagctctaagta5640ttaccagcgcttcacccccgaaatcgtcgcacgaccgaaggtgagagggcaggccgggag5700aatgaactactactggaccctgctggatcaaggcgacactattaccttcgaggctaccgg5760caacttgatcgccccgtggcacgcgttcgccctcaataaaggatctaatagcggcataat5820gatgagtgatgcccacgtgcataactgcaccacgaagtgccagacccctcacggcgcact5880gaaaagcaatctgccctttcagaatgtgcaccccatcaccatcggcgagtgccccaagta5940tgttaaaagcactcagctccgcatggccaccggactgcgcaacatcccgagcatccaatc6000ccgcggactgttcggcgcaatcgcgggctttatagagggcggctggaccggcatgatcga6060cggctggtacggctaccaccatcaaaatgagcaaggttccggctacgccgcagaccagaa6120gagcacccaaatagcaatcgatggcatctccaacaaggtgaacagcgtgatcgaaaagat6180gaacatccagttcacaagcgtggggaaggagttcaataacctggaaaagcgcatcgagaa6240tctgaacaagaaggttgacgatgggttcctcgatgtctggacctataacgccgagctcct6300gatactgcttgagaacgagcgcaccctggacttccacgacttcaacgtgaaaaacctgta6360cgaaaaggtcaagtcacagttgcgaaacaatgcgaaggagataggcaacggctgcttcga6420gttctatcacaagtgtgacaacgagtgcatggagagcgtcaagaacggcacttacaacta6480cccgcgctactctgaggagagtaagctcaaccgcgaagagattgacggcgtgaaactgga6540aagcgttggtgtccatcagatcctggccatctacagcaccgtggctagctctctggttct6600gttggtgagcctgggcgctataagcttttggatgtgttctaatgggagcctgcagtgccg6660catctgcatctgaggtac6678<210>26<211>566<212>prt<213>influenzaavirus<400>26metlysalaileleuvalvalleuleutyrthrphealathralaasn151015alaaspthrleucysileglytyrhisalaasnasnserthraspthr202530valaspthrvalleuglulysasnvalthrvalthrhisservalasn354045leuleugluasplyshisasnglylysleucyslysleuargglyval505560alaproleuhisleuglylyscysasnilealaglytrpileleugly65707580asnproglucysgluserleuserthralasersertrpsertyrile859095valgluthrserserseraspasnglythrcystyrproglyaspphe100105110ileasptyrglugluleuarggluglnleuserservalserserphe115120125gluargphegluilepheprolysthrsersertrpproasnhisasp130135140serasnlysglyvalthralaalacysprohisalaglyalalysser145150155160phetyrlysasnleuiletrpleuvallyslysglyasnsertyrpro165170175lysleuserlyssertyrileasnasplysglylysgluvalleuval180185190leutrpglyilehishisproserthrseralaaspglnglnserleu195200205tyrglnasnalaaspalatyrvalphevalglythrserargtyrser210215220lyslysphelysprogluilealaileargprolysvalargaspgln225230235240gluglyargmetasntyrtyrtrpthrleuvalgluproglyasplys245250255ilethrpheglualathrglyasnleuvalvalproargtyralaphe260265270alametgluarglysalaglyserglyileileileseraspthrpro275280285valhisaspcysasnthrthrcysglnthrprolysglyalaileasn290295300thrserleupropheglnasnilehisproilethrileglylyscys305310315320prolystyrvallysserthrlysleuargleualathrglyleuarg325330335asnileproserileglnserargglyleupheglyalailealagly340345350pheilegluglyglytrpthrglymetvalaspglytrptyrglytyr355360365hishisglnasngluglnglyserglytyralaalaaspleulysser370375380thrglnasnalaileaspgluilethrasnlysvalasnservalile385390395400glulysmetasnthrglnphethralavalglylysglupheasnhis405410415leuglulysargilegluasnleuasnlyslysvalaspaspglyphe420425430leuaspiletrpthrtyrasnalagluleuleuvalleuleugluasn435440445gluargthrleuasptyrhisaspserasnvallysasnleutyrglu450455460lysvalargserglnleulysasnasnalalysgluileglyasngly465470475480cysphegluphetyrhislyscysaspasnthrcysmetgluserval485490495lysasnglythrtyrasptyrprolystyrsergluglualalysleu500505510asnargglugluileaspglyvallysleugluserthrargiletyr515520525glnileleualailetyrserthrvalalaserserleuvalleuile530535540valserleuglyalaileserphetrpmetcysserasnglyserleu545550555560glncysargilecysile565<210>27<211>566<212>prt<213>influenzaavirus<400>27metlysthrvalilealaleusertyrilephecysleuvalphegly151015glnaspleuproglylysglyasnasnthralathrleucysleugly202530hishisalavalproasnglythrleuvallysthrilethraspasp354045glnilegluvalthrasnalathrgluleuvalglnasnphesermet505560glylysilecysasnasnprohisargileleuaspglyalaasncys65707580thrleuileaspserleuleuglyaspprohiscysaspglyphegln859095asnglulystrpaspleupheilegluargserlysalapheserasn100105110cystyrprotyraspvalproglutyrthrserleuargserleuile115120125alaserserglythrleugluphethrasngluasnpheasntrpthr130135140glyvalthrglnasnglyglyserseralacyslysargglyproasn145150155160asnserphepheserlysleuasntrpleutyrlysserglyasnthr165170175tyrprometleuasnvalthrmetproasnseraspasppheasplys180185190leutyriletrpglyvalhishisproserthrasparggluglnile195200205asnleutyrvalglnalaserglylysileilevalserthrlysarg210215220serglnglnthrileileproasnileglyserargprotrpvalarg225230235240glyleuserserargileseriletyrtrpthrilevallysprogly245250255aspileleuileileasnserasnglyasnleuilealaproarggly260265270tyrphelysmetglnthrglylysserservalmetargseraspala275280285proileaspthrcysasnserglucysilethrproasnglyserile290295300proasnasplyspropheglnasnvalasnlysilethrtyrglyala305310315320cysprolystyrilelysglnasnthrleulysleualathrglymet325330335argasnileprogluargglnthrargglyilepheglyalaileala340345350glypheilegluasnglytrpgluglymetvalasnglytrptyrgly355360365phearghisglnasnsergluglyileglyglnalaalaaspleulys370375380serthrglnalaalaileasnglnileasnglylysleuasnargval385390395400ileglulysthrasnglulysphehisglnileglulysglupheser405410415gluvalgluglyargileglnaspleugluargtyrvalgluaspthr420425430lysileaspleutrpsertyrasnalagluleuleuvalalaleuglu435440445asnglnhisthrileaspleuthraspserglumetasnlysleuphe450455460glulysthrarglysglnleuarggluasnalagluaspmetglyasn465470475480glycysphelysiletyrhislyscysaspasnsercysmetgluser485490495ileargasnglythrtyrasphisasnglutyrargaspglualaval500505510asnasnargpheglnilelysservalgluleulysserglytyrlys515520525asptrpileleutrpileserphealailesercyspheleuleucys530535540alavaltrpleuglypheilemettrpalacysglnlysglyasnile545550555560argcysasnilecysile565<210>28<211>566<212>prt<213>influenzaavirus<400>28metglualalysleuphevalleuphecysalaphethralaleulys151015alaaspthrilecysvalglytyrhisalaasnasnserthraspthr202530valaspthrileleuglulysasnvalthrvalthrhisservalasn354045leuleugluasnserhisasnglylysleucysserleuasnglylys505560alaproleuglnleuglyasncysasnvalalaglytrpileleugly65707580asnproglucysaspleuleuleuthralaasnsertrpsertyrile859095ilegluthrserasnserlysasnglyalacystyrproglygluphe100105110alaasptyrglugluleuarggluglnleuserthrvalserserphe115120125gluargphegluilepheprolysalathrsertrpproasnhisasp130135140thrthrlysglythrthrvalsercysserhisserglyalaasnser145150155160phetyrargasnleuleutrpilevallyslysglyasnsertyrpro165170175lysleuserlyssertyrthrasnasnlysglylysgluvalleuval180185190iletrpglyvalhishisproprothraspseraspglnglnthrleu195200205tyrglnasnasnhisthrtyrvalservalglyserserlystyrtyr210215220glnargphethrprogluilevalalaargprolysvalargglygln225230235240alaglyargmetasntyrtyrtrpthrleuleuaspglnglyaspthr245250255ilethrpheglualathrglyasnleuilealaprotrphisalaphe260265270alaleuasnlysglyserasnserglyilemetmetseraspalahis275280285valhisasncysthrthrlyscysglnthrprohisglyalaleulys290295300serasnleupropheglnasnvalhisproilethrileglyglucys305310315320prolystyrvallysserthrglnleuargmetalathrglyleuarg325330335asnileproserileglnserargglyleupheglyalailealagly340345350pheilegluglyglytrpthrglymetileaspglytrptyrglytyr355360365hishisglnasngluglnglyserglytyralaalaaspglnlysser370375380thrglnilealaileaspglyileserasnlysvalasnservalile385390395400glulysmetasnileglnphethrservalglylysglupheasnasn405410415leuglulysargilegluasnleuasnlyslysvalaspaspglyphe420425430leuaspvaltrpthrtyrasnalagluleuleuileleuleugluasn435440445gluargthrleuaspphehisasppheasnvallysasnleutyrglu450455460lysvallysserglnleuargasnasnalalysgluileglyasngly465470475480cysphegluphetyrhislyscysaspasnglucysmetgluserval485490495lysasnglythrtyrasntyrproargtyrserglugluserlysleu500505510asnargglugluileaspglyvallysleugluservalglyvalhis515520525glnileleualailetyrserthrvalalaserserleuvalleuleu530535540valserleuglyalaileserphetrpmetcysserasnglyserleu545550555560glncysargilecysile565<210>29<211>564<212>prt<213>influenzaavirus<400>29metlysalalysleuleuileleutrpcysalaleuseralathrasp151015alaaspthrilecysileglytyrhisalaasnasnserthraspthr202530valaspthrvalleuglulysasnvalthrvalthrhisservalasn354045leuleugluaspasnhisasnglylysleucyslysleulysglyval505560alaproleuglnleuglylyscysserilealaglytrpileleugly65707580asnproglucysgluserleupheserlyslyssertrpsertyrile859095alagluthrproasnalagluasnglyilecystyrproglytyrphe100105110serasptyrglugluleuarggluglnleuserservalserserphe115120125gluargphegluilepheprolysglusersertrpprolyshisser130135140ileglyalathralasercysserlysglnglyargserserphetyr145150155160argasnleuleutrpleuthrglulysasnglysertyrproasnleu165170175serlyssertyrvalasnasplysgluarggluvalleuvalleutrp180185190glyvalhishisproserasnilegluaspglnargalailetyrarg195200205lysgluthralatyrvalservalmetserserleutyrasnargarg210215220phethrprogluilealalysargprolysileargasnglnglugly225230235240argileasntyrtyrtrpthrleuleugluprolysaspthrileile245250255pheglualaasnglyasnleuilealaprotrptyralaphealaleu260265270serargglyphegluserglyileilevalserasnalasermetasp275280285glucysaspalalyscysglnthrproglnglyalaileasnserser290295300leupropheglnasnvalhisprovalthrileglyglucysprolys305310315320tyrvallysserthrlysleulysmetalathrglyleuargasnile325330335proserileglnthrargglyleupheglyalailealaglypheile340345350gluglyglytrpthrglymetileaspglytrptyrglytyrhishis355360365glnasngluglnglyserglytyralaalaaspglnlysserthrgln370375380asnalaileasnglyilethrasnlysvalasnservalileasplys385390395400metasnthrglnphethralavalglylysglupheasnlysleuglu405410415lysargmetgluasnleuasnlyslysvalaspaspglypheleuasp420425430iletrpthrtyrasnalagluleuleuvalleuleugluasngluarg435440445thrleuaspphehisaspserasnvallysserleutyrglulysval450455460lysglyglnleulysasnasnalalysgluileglyasnglycysphe465470475480gluphetyrhislyscysasnasnglucysmetaspservallysasn485490495glythrtyrasptyrproargtyrserglugluserlysleuasnarg500505510glulysileaspglyvalgluleulyssermetglyvaltyrglnile515520525leualailetyrserthrvalalaserserleuvalleuleuvalser530535540leuglyalathrserphetrpmetcysserasnglyserleuglncys545550555560argilecysile<210>30<211>19<212>prt<213>artificialsequence<220><223>gslinker<400>30hismetglyglyserglyglyglyglyserglyglyglyglysergly151015glyglythr<210>31<211>947<212>dna<213>artificialsequence<220><223>synthesizeddnasequenceoftruncatedglycoproteinc(gc)includingrestrictionsitesforsubcloning<400>31gcggccgcatgaaggcgatcctggttgtgctgctgtacacctttgccaccgccaacgccg60atacgctgatcaactgcaagaacatccagagcacccagctgacaatcgagcacctgagca120agtgcatggccttctaccagaacaagaccagcagccccgtcgtgatcaacgagatcatct180ccgacgccagcgtggacgaacaggaactgattaagtctctgaacctgaactgcaacgtga240tcgaccggttcatcagcgagtccagcgtgatcgagacacaggtgtactacgagtatatca300agagccagctgtgtccactgcaagtgcacgatatcttcaccatcaacagcgccagcaaca360tccagtggaaggccctggcccgcagctttaccctgggcgtgtgcaacaccaacccccaca420agcacatctgccggtgcctggaatccatgcagatgtgtaccagcaccaagaccgaccacg480ccagagagatgagcatctactacgacggccaccccgacagattcgagcacgacatgaaga540ttatcctgaatatcatgcggtacatcgtgcccggcctgggcagagtgctgctggaccaga600tcaagcagaccaaggactaccaggccctgagacacatccagggcaagctgagccccaagt660cccagagcaacctgcagctgaagggcttcctggaattcgtggacttcatcctgggcgcca720acgtgaccattgagaaaaccccccagaccctgaccaccctgagcctgattcatatgggag780gttccggaggtggaggttccggaggtggaggttccggaggtggcaccatactggccattt840acagcacagttgcgagcagcctggtcctgatcgtgagcctgggtgctatatcattctgga900tgtgcagcaacggctctctccagtgccgcatctgtatctgaggtacc947<210>32<211>3060<212>dna<213>artificialsequence<220><223>dnafragmentusedforredrecombinationtogenerateprach-se-70-455-sbvgc,<400>32tctagactcgagcgcaagccctacacgcgctacccctgctttcaacgcgtcaacctgcac60attgacggggagtttctggttcacaagatgctagcgttcaatgccgcgatgcgcccatcg120gccgaggagctgctgtcatacccaatgtttgctcaactttaggatgactaacctgtttct180gggaggagacagcgtgggcgacggtgtataaagttggtctgctttcaagccctgccactg240cgctacagtgccaccaactgtaaagcggtagtaagctgcagtggtcgactggtggtagca300tatactaccttatttatacgctccgagctgtttttcagcatgctagcacccaacgccgag360cgagagtatataactcccatcattgcccacaagcttatgccacttattagcgtccgctct420gccgtttgcttagtcataatatctaccgccgtttacgcagcagacgctatctgcgacaca480attggatttgcgataccgcgcatgtggatgtgtattttaatgagatcaacctccatgaag540cgtaactagggggcctcccactgaggcactaccggcttagcagctgactaacacagtata600aaacgtgagaagaaatcagtctcatgcgccattagcgctaggctagttagcgtggaggac660cggagcgctaccgccagcagtttcatccgcctggttacgggtttgttaacacctaccggt720gttttaccgctaccataggatccgatccatgggcggccgcatgaaggcgatcctggttgt780gctgctgtacacctttgccaccgccaacgccgatacgctgatcaactgcaagaacatcca840gagcacccagctgacaatcgagcacctgagcaagtgcatggccttctaccagaacaagac900cagcagccccgtcgtgatcaacgagatcatctccgacgccagcgtggacgaacaggaact960gattaagtctctgaacctgaactgcaacgtgatcgaccggttcatcagcgagtccagcgt1020gatcgagacacaggtgtactacgagtatatcaagagccagctgtgtccactgcaagtgca1080cgatatcttcaccatcaacagcgccagcaacatccagtggaaggccctggcccgcagctt1140taccctgggcgtgtgcaacaccaacccccacaagcacatctgccggtgcctggaatccat1200gcagatgtgtaccagcaccaagaccgaccacgccagagagatgagcatctactacgacgg1260ccaccccgacagattcgagcacgacatgaagattatcctgaatatcatgcggtacatcgt1320gcccggcctgggcagagtgctgctggaccagatcaagcagaccaaggactaccaggccct1380gagacacatccagggcaagctgagccccaagtcccagagcaacctgcagctgaagggctt1440cctggaattcgtggacttcatcctgggcgccaacgtgaccattgagaaaaccccccagac1500cctgaccaccctgagcctgattcatatgggaggttccggaggtggaggttccggaggtgg1560aggttccggaggtggcaccatactggccatttacagcacagttgcgagcagcctggtcct1620gatcgtgagcctgggtgctatatcattctggatgtgcagcaacggctctctccagtgccg1680catctgtatctgaggtaccaataaacgcggtatgtctaccttcaagcctatgatgaacgg1740atgtttggtgtttgcggctattataacgctcttgagttttatgctatctctgggaacatg1800cgaaaattacaggcgtgtggttcgggatcctagggataacagggtaatcgatttattcaa1860caaagccacgttgtgtctcaaaatctctgatgttacattgcacaagataaaaatatatca1920tcatgaacaataaaactgtctgcttacataaacagtaatacaaggggtgttatgagccat1980attcaacgggaaacgtcttgctcgaggccgcgattaaattccaacatggatgctgattta2040tatgggtataaatgggctcgcgataatgtcgggcaatcaggtgcgacaatctatcgattg2100tatgggaagcccgatgcgccagagttgtttctgaaacatggcaaaggtagcgttgccaat2160gatgttacagatgagatggtcagactaaactggctgacggaatttatgcctcttccgacc2220atcaagcattttatccgtactcctgatgatgcatggttactcaccactgcgatccccggg2280aaaacagcattccaggtattagaagaatatcctgattcaggtgaaaatattgttgatgcg2340ctggcagtgttcctgcgccggttgcattcgattcctgtttgtaattgtccttttaacagc2400gatcgcgtatttcgtctcgctcaggcgcaatcacgaatgaataacggtttggttgatgcg2460agtgattttgatgacgagcgtaatggctggcctgttgaacaagtctggaaagaaatgcat2520aagcttttgccattctcaccggattcagtcgtcactcatggtgatttctcacttgataac2580cttatttttgacgaggggaaattaataggttgtattgatgttggacgagtcggaatcgca2640gaccgataccaggatcttgccatcctatggaactgcctcggtgagttttctccttcatta2700cagaaacggctttttcaaaaatatggtattgataatcctgatatgaataaattgcagttt2760catttgatgctcgatgagtttttctaaaataaacgcggtatgtctaccttcaagcctatg2820atgaacggatgtttggtgtttgcggctattataacgctcttgagttttatgctatctctg2880ggaacatgcgaaaattacaggcgtgtggttcgggatccgaccctgttggtgggtgcggtt2940ggactcagaatcttggcgcaggcatggaagtttgtcggtgacgaaacatacgacaccatc3000cgcgcagaagcaaagaatttagagacccacgtaccctcaagtgctgcagagtcgtctaga3060<210>33<211>17<212>dna<213>artificialsequence<220><223>primer<400>33cgtgcgcggatacatcg17<210>34<211>16<212>dna<213>artificialsequence<220><223>primer<400>34cgcttcgcaggtgggc16<210>35<211>19<212>dna<213>artificialsequence<220><223>primer<400>35gactggtggtagcatatac19<210>36<211>19<212>dna<213>artificialsequence<220><223>primer<400>36gatcaacgagatcatctcc19<210>37<211>17<212>dna<213>artificialsequence<220><223>primer<400>37ctggagagagccgttgc17当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 一种基于转录组测序构建的棒状杆菌组成型表达载体启动子的制造方法与工艺
- 一种棒状杆菌组成型表达载体启动子、含有该启动子及gnd基因的表达载体的制造方法与工艺
- 一种棒状杆菌组成型表达载体启动子、含有该启动子及lrp基因的表达载体的制造方法与工艺
- 一株呕吐毒素降解菌及其应用的制造方法与工艺
- 一种具有家蚕血细胞特异性活性的启动子的鉴定方法与流程
- 一种改造的质粒复制子及其应用的制造方法与工艺
- 一种水稻种子特异性启动子Posseed及其应用的制造方法与工艺
- 一种在水稻幼穗特异表达的启动子及其应用的制造方法与工艺
- 一种表达外源基因的启动子及其应用的制造方法与工艺
- 可诱导的CRISPRon或CRISPRi小鼠胚胎干细胞及其应用的制造方法与工艺