一种高效表达P53抑癌蛋白的T细胞、制备方法及应用与流程
本发明涉及分子细胞生物学领域,特别涉及p53抑癌蛋白的应用和转化,以及用p53抑癌蛋白治疗血液和实体瘤的免疫疗法。
背景技术:
p53蛋白是1979年由davidlane发现并根据其分子量大小命名的,它是核蛋白四聚体,通过几种转录后修饰作用,如磷酸化、乙酰化、泛素化、类泛素化、甲基化等修饰,可以跟多种蛋白质及dna相互作用[1],影响精细、复杂的细胞调节过程。多种信号可以诱导p53激活,如dna损伤、营养饥饿、热休克、病毒感染、ph改变、缺氧和原癌基因激活等。特别地,p53作为转录因子,可以通过特异性绑定p53-效应元件(p53res)激活或者抑制基因表达[2]。p53位于17号染色体的短臂上(17p13.1)[3],含有11个外显子,10个内含子。其中除1号外显子为无意编码外,2-11号外显子编码393个氨基酸的蛋白。p53氨基酸序列中,93-292氨基酸区域是dna结合区域以及人类肿瘤中的突变高发生区[4]。95%的p53碱基置换突变发生在氨基酸保守序列上(密码子117–142、171–181、234–258和270–286)[5,6]。p53蛋白的n-端富含大量的氨基酸残基,可以作为转录激活因子[7];c-端寡聚区,包含dna非特异性结合区域,可以识别并结合损伤的dna[8]。如果p53基因错意突变发生在特定的n-或者c-端区域,将会影响基因抑制肿瘤的功能。
p53基因是迄今为止已发现的与人类肿瘤发生相关性最高的抑癌基因,但是这种基因的突变存在着致瘤性转化[9]。据报道,超过50%人类恶性肿瘤的发生是由于p53基因突变引起的,超过90%p53基因点突变发生在中央核心结构域上[10]。其中约73%突变属于由错义突变引起的蛋白构象的改变[11,12]。一方面,发生点突变的p53对肿瘤相关信号通路的调节失活,转而易于发生癌变;另一方面,即使p53不存在突变,但是p53参与的信号通路被干扰依然可以使人体罹患癌症[13]。卵巢癌中p53基因突变率最高,主要为点突变,29%~74%。okamoto等报道,80%卵巢癌有p53等位基因丢失,29%卵巢癌有p53基因突变。子宫内膜癌中,32%的病人伴有p53等位基因丢失。宫颈癌中的突变率较低,约3%~7%。另外,70%~80%肺癌患者也存在p53基因突变。通过免疫组化等手段可以检测到突变的p53蛋白大量聚集在细胞核内[14,15]。
随后研究发现,p53不仅具有抑制作用,同时还可以维持各种细胞功能的发挥,因而被称作“基因组的守护者”[16,17]。现在认为抑癌基因的突变相比较于癌基因的突变对于癌症的发生更加危险。通过使用适当的肿瘤抑制基因或去除掉致癌的相应癌基因是诱导癌细胞死亡的一种很有希望的抗肿瘤治疗策略。外源性p53基因的异位表达能有效的诱导许多p53失活的肿瘤细胞的死亡。为了诱导外源性p53基因的表达,在许多种癌症的临床前体外和体内实验中经常使用腺病毒载体来表达p53。这些癌症包括:非小细胞肺癌,头颈癌,恶性脑肿瘤,卵巢癌,膀胱癌,子宫颈癌,结肠直肠癌以及食管癌等。
在包括人在内的机体中普遍存在癌基因和抑癌基因,它们的功能障碍能够导致癌症的发生和发展。现在认为抑癌基因的突变相比较于癌基因的突变对于癌症的发生更加危险。通过使用适当的肿瘤抑制基因或去除掉致癌的相应癌基因是诱导癌细胞死亡的一种很有希望的抗肿瘤治疗策略[6,18]。外源性p53基因的异位表达能有效的诱导许多p53失活的肿瘤细胞的死亡。为了诱导外源性p53基因的表达,在许多种癌症的临床前体外和体内实验中经常使用腺病毒载体来表达p53。这些癌症包括[15,19,20]:非小细胞肺癌,头颈癌,恶性脑肿瘤,卵巢癌,膀胱癌,子宫颈癌,结肠直肠癌以及食管癌等。
腺病毒-p53(ad-p53)载体介导的癌症基因治疗已经应用于临床治疗。临床前体外和体内实验的研究表明,ad-p53-介导的基因治疗能在肿瘤细胞中诱导广泛的抗肿瘤效应,并且在正常细胞中具有低细胞毒性。携带p53功能损伤的肿瘤细胞通常对传统的化疗放疗引起的遗传毒性压力具有很强的耐受性[21,22]。研究表明腺病毒介导的p53野生型基因表达能增强这些肿瘤细胞对传统的化疗和放疗的敏感性。最近,在多种肿瘤病人的临床实验中,已经对ad-p53载体作为单一疗法或与传统的化疗和放疗结合的组合疗法的效果进行了评估。尽管临床研究表明在各种肿瘤患者中,复制缺陷型ad-p53载体是安全可行的,并且具有良好的耐受性,但是通过这种ad-p53载体不可能在每一个肿瘤细胞中诱导深刻的外源性p53蛋白表达。为了提高治疗晚期肿瘤患者的效果,ad-p53载体较低的转基因效率是一个必须克服的主要问题。另一方面,市面上的ad-p53针对实体瘤的治疗主要是通过瘤体注射,操作复杂且有一定的危险性。探索一种既能有效高表达p53又能简单操作的肿瘤治疗方法具有更广阔的前景。
申请号02815303.0,发明名称为与p53结合的t细胞受体分子以及其用途的发明专利中提供了一种t细胞受体(tcr)分子,它可以与衍生自p53蛋白质的肽结合,特别是人类p53蛋白质。该tcr分子包含杂二聚分子以及单链分子,且该tcr分子特异性地结合至显现于上下文的hla分子系统,特别是hla-a2.1中的p53蛋白质的一个序列段,尤其是氨基酸264-273的位置。该发明还揭示了制造及使用这类tcr分子的方法。该发明具有广泛的用途,包括治疗用途以及检测表达p53蛋白质的细胞的用途。
但是该发明专利仅仅表达了p53蛋白序列的表位,不具有一定的空间构象;另外,该发明只用于识别p53264-273的位置,而非p53蛋白的全氨基酸序列。该发明可以用于tcr对符合264-273肽段的识别,用于结合包含该肽段的p53或者鉴定含有该肽段的p53,不以表达p53全蛋白为目的。
技术实现要素:
针对以上技术缺陷,本发明建立高效表达p53的载体体系,以免疫t细胞为表达载体,可以高效表达不同亚型p53,更有效地发挥p53的抗肿瘤活性。
本发明的第一目的旨在构建高表达p53的载体,进一步制备高效表达p53抑癌蛋白的t细胞。
因此,本发明提供一种高效表达p53抑癌蛋白的t细胞,由p53真核表达载体转染t细胞得到;所述p53真核表达载体,是将p53基因连接至pcdna3.1载体中构建而成。
本发明的第二目的在于提供所述高效表达p53抑癌蛋白的t细胞的制备方法。本发明高效表达p53抑癌蛋白的t细胞的制备方法,步骤为:
(1)构建稳定表达p53真核表达载体:将p53基因与pcdna3.1载体连接,构建p53真核表达载体pcdna3.1-p53;
(2)转染免疫t细胞:将新鲜的t细胞加入含有10%fbs的rpmi1640完全培养基中,用抗cd3、cd28抗体、il-2刺激t细胞,第三天用lipofectamine2000将构建好的pcdna3.1-p53载体转染t细胞,筛选得到稳定过表达p53的t细胞株p53-t细胞。
本发明制备的t细胞,可以-80℃冻存,长期保存。使用时,复苏细胞,并在体外大量扩增,使其分别达到108数量级,通过静脉注射单独或者与其他辅助成分共同输入病人体内,达到缓解癌症病人病症的作用。
因此,本发明还提供所述高效表达p53抑癌蛋白的t细胞在制备用于治疗肿瘤的药物方面的应用。
所述肿瘤包括:肝癌、非小细胞肺癌、头颈癌、恶性脑肿瘤、卵巢癌、膀胱癌、子宫颈癌、结肠直肠癌以及食管癌。
本发明具有如下有益效果:
(1)t细胞既作为p53的生产细胞,同时又是可以杀死肿瘤细胞的免疫细胞,具有广谱治疗癌症的作用。
(2)大量免疫细胞t细胞输到病人体内后,能增进病人的免疫能力,刺激并激活病人的免疫系统,从而进一步增强病人自身对癌细胞的杀伤能力。
(3)t细胞产生的p53蛋白能分泌到病人血液中,并随血液循环到达身体各处,因此不仅能杀死局部的实体肿瘤,并且也具有杀死已经转移的肿瘤细胞的作用。
(4)本发明可以表达不同亚型的p53,达到针对个体不同症状治疗的效果。p53基因不仅在不同的肿瘤中表达不同的亚型,并且不同的亚型也存在不同的转录活性和肿瘤抑制因子功能。由于p53亚型表达的重要性与复杂性,因此本发明以t细胞为载体,生产含有天然的、不同构象的p53全蛋白,用于肿瘤患者的治疗,疗效更为确切。
附图说明
图1为本发明构建的p53真核表达载体结构示意图。
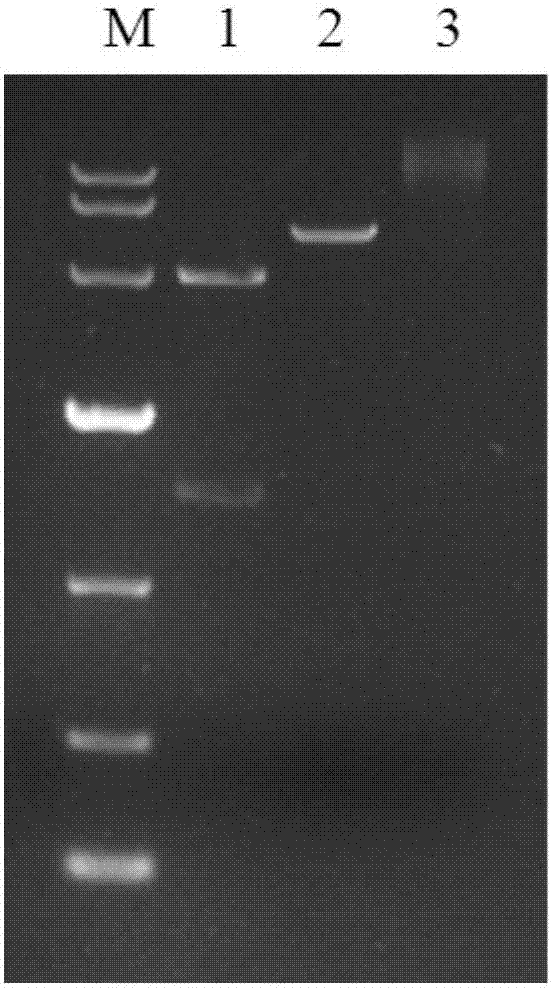
图2为表达载体pcdna3.1-p53琼脂糖凝胶电泳图;其中1,bamhi酶切载体;2,noti酶切载体;3,对照组。
图3为westernblotting检测p53过表达载体在t细胞中的表达情况。
具体实施方式
下面结合具体试验方法对本发明的技术方案及其所产生的技术效果做进一步的阐述。应当理解,下述说明仅是为了解释本发明,但不以任何方式对本发明加以限制,基于本发明所作的任何变换或替换,均属于本发明的保护范围。本发明所述方法如无特殊说明,均为本领域常规方法。
实施例1.高效表达p53抑癌蛋白的t细胞的制备方法
1.真核细胞表达载体的构建
1-1回收目的基因
bamhi-p53-bamhi基因合成由金斯瑞生物科技有限公司合成,如图1。得到的含有目的基因的载体通过bamhi酶切后回收基因片段。目的基因酶切体系如下:
该酶切体系置于37℃水浴锅中,酶切过夜。酶切产物用快速dna产物纯化试剂盒纯化。该过程遵照试剂盒说明书进行。
1-2目的基因与载体连接
pcdna3.1(+)空载体经bamhi酶切回收后去磷酸化处理。用dna连接试剂盒ligationhighver.2将回收的p53片段与酶切过的pcdna3.1(+)空载连接,16℃水浴过夜。
1-3连接载体转化
将16μl上述连接产物全部加入100μl大肠杆菌dh5α感受态细胞中,冰浴30min,立即在42℃水浴中热击90s后移至冰上冰浴2min。加入400μllb培养基,37℃,200rpm培养1h。滴加10μl菌液至含有100μg·ml-1氨苄青霉素的lb固体平板上,均匀涂布。培养皿于37℃培养箱中倒置培养约16h。
1-4阳性菌落筛选
挑选大肠杆菌单克隆,标记号码后加入4ml含有氨苄青霉素的lb液体培养基中培养过夜。次日取2ml菌液,用质粒小提试剂盒(康维世纪)提取转化质粒。琼脂糖凝胶电泳检测结果。对照组为未酶切相应质粒,如图2。测序比对正确的阳性克隆菌为含有pcdna3.1-p53载体的质粒。
2.稳定表达p53蛋白的t细胞的筛选
将新鲜的t细胞加入含有10%fbs的rpmi1640完全培养基中,用抗cd3、cd28抗体、il-2刺激t细胞,第三天用lipofectamine2000将构建好的pcdna3.1-p53载体转染细胞。筛选稳定过表达p53的t细胞株,p53-t细胞,并用于westernblotting检测。
3.westernblotting检测p53过表达载体在t细胞中的表达
3-1westernblot相关试剂
(1)5×sds-page电泳缓冲液:称取tris15.1g,sds5.0g,甘氨酸94g,去离子水定容至1l备用。工作浓度为1×。
(2)5×sds-page上样缓冲液:称取sds0.5g,溴酚蓝25mg,加入1mtris-hcl(ph6.8)1.25ml,甘油2.5ml,去离子水定容至5ml,保存备用。
(3)膜转移缓冲液:称取甘氨酸2.9g,tris5.8g,sds0.37g,适量去离子水溶解后加入甲醇200ml,去离子水定容至1l后4℃保存备用。
(4)tbst缓冲液:称取nacl8.8g,加入1mtris-hcl(ph8.0)20ml后去离子水定容1l,加入吐温200.5ml,混匀备用。
(5)5%脱脂牛奶封闭液:用tbst配制即可。
(6)ripa裂解液:150mmnacl,5mmedta,5mm焦磷酸钠,25mmtris(ph7.4),1%tritonx-100,0.1%sds,0.5%脱氧胆酸钠。使用时添加1%pmsf,0.1%抑肽酶。
3-2总蛋白的提取
(1)取步骤2中的细胞培养皿置于冰上,弃培养液。预冷的pbs清洗3次后将残留液体吸干净。加入预冷的ripa裂解液(700μl/10-cm培养皿;100μl/6孔板)后,取细胞刮铲将贴壁细胞刮下,冰浴30min。收集裂解液涡旋震荡,12000rpm,4℃离心15min。小心吸取上清至干净的1.5ml离心管中,置于冰上。取一部分蛋白液用于测定蛋白浓度,剩余蛋白液加入5×上样缓冲液(终浓度为1×),混匀后煮沸5min。得到的蛋白-20℃冻存备用。对照组为不加入载体转染的t细胞。
(2)蛋白电泳:待测蛋白上样至聚丙烯酰胺凝胶蛋白点样孔中,加电泳缓冲液,80v,30min,之后设置160v至溴酚蓝到达分离胶底部。
(3)转膜:取与浸泡于转膜缓冲液中分离胶大小相同的滤纸8片,pvdf膜1片。使用前pvdf膜置于甲醇溶液中浸泡10s。转膜仪器使用trans-blotturbo蛋白转印系统。在转膜仪下电极板上由下至上贴沾有转膜缓冲液的滤纸片、pvdf膜、分离胶、滤纸片。注意操作过程中排除转膜系统中的气泡。转膜条件:25v恒压,5-10min(根据转膜蛋白分子量确定)。
(4)封闭:将印有蛋白的pvdf膜置于5%脱脂牛奶中,37℃孵育2hr。
(5)抗体孵育:封闭后pvdf膜置于tbst缓冲液中,遵照说明书使用浓度加入一抗:anti-p53(abcam,ab1101,43.6kda),4℃,慢摇孵育过夜。tbst清洗pvdf膜3次,10min/次。加入相应二抗,37℃慢摇孵育1hr,重复以上步骤,tbst中清洗3次。
(6)显色:遵照说明书配制pro-lighthrp化学发光检测试剂,并滴加至待显色pvdf膜上,用chemidoctmxrs+成像系统曝光采集图像。结果如图3所示。
实验例1.p53-t细胞对smmc-7721肝癌细胞的体外杀伤实验
冻存的p53-t细胞解冻复苏后,加入10%fbs的rpmi1640完全培养基中,用抗cd3、cd28抗体、il-2刺激t细胞,培养3天后,换液,补充营养因子继续扩大培养。第五天,加入肿瘤抗原hsp70培养24小时后备用(对照组不加抗原),同时,将培养至指数期的smmc-7721细胞,胰酶消化后铺板96孔板,5000个细胞/孔。24小时后,将准备好的p53-t细胞加入铺板的smmc-7721细胞中。效靶比50:1,实验分组见表1,每组平行实验5次。细胞共培养24小时后,加入mtt,20μl/孔,继续培养6小时后,吸去上清,每孔加入150μldmso,震荡混匀,酶标仪492nm处测od值,记录。按以下公式计算p53-t细胞对smmc-7721细胞的体外杀伤活性:
杀伤活性=100-((实验组od值-效应细胞od值)/靶细胞od值×100%)
采用t检验对数据进行分。分析结果见表1。
表1p53-t细胞对smmc-7721肝癌细胞体外杀伤活性分析
表1结果表明,与单纯t细胞杀伤活性相比较,p53-t的肿瘤杀伤活性提高了12.5个百分点。由于,本发明以t细胞为载体,产生含有天然的、不同构象的p53全蛋白,因此,用于肿瘤患者的治疗,疗效更为确切。
讨论:1984年,matlashewski第一次发现了p53的亚型[23]。一年后,rotter等研究了小鼠p53c-端选择性剪切后的变体,并很快在人类细胞中得到证实[24]。根据c-端(α,β,γ)和n-端的不同(△40p53,△133p53,△160p53),理论上,p53通过转录后修饰可以翻译成12种亚型蛋白(p53α,p53β,p53γ,△40p53α,△40p53β,△40p53γ,△133p53α,△133p53β,△133p53γ,△160p53α,△160p53β,和△160p53γ)[18]。p53基因不仅在不同的肿瘤中表达不同的亚型,并且不同的亚型也存在不同的转录活性和肿瘤抑制因子功能,这种交错复杂的结构,共同影响着细胞的生物学功能。由于p53亚型表达的重要性与复杂性,已经有很多学者致力于p53不同亚型的临床研究,试图在mrna和蛋白水平上阐述不同p53的亚型与肿瘤类型的相关性[25]。fujita等研究发现,可以通过报道p53β和△133p53α的不同比例,预测结直肠癌腺瘤到恶性肿瘤的发展等级[25]。肾细胞癌中,p53β的mrna和蛋白表达水平增高预示着癌症的发展进程[26]。黏蛋白卵巢癌中△40p53α的表达,提高了病人无复发情况下的存活率[27]。对30例原发性乳腺癌的研究发现,其中18例伴随p53β和p53γ的表达缺失[28];12例存在△133p53的过表达[28]。胆管癌中,△133p53的表达具有不良预后[29]。急性髓细胞白血病中p53β和p53γ的过表达是化疗后的病情好转的重要指标[30]。综合报道结果证实,不同亚型的p53在不同类型肿瘤中的表达指示了肿瘤的分级、临床反应和预后情况。
研究表明,通过过量表达p53蛋白可以有效的诱导多种肿瘤细胞的死亡[31]。迄今,虽然在p53靶向治疗癌症方面取得了很大的进展,但是,目前在有效治疗人类癌症方面还存在巨大的不足。为了更好的治疗癌症,减轻病人的痛苦,并最终达到根治癌症的目的,更为有效的p53抑癌基因肿瘤治疗策略亟待进一步研发。
基于以上分析,我们设计研发了新的更有效地基于人抑癌基因p53的靶向治疗肿瘤的策略。从其对肿瘤杀伤性实验中可以看出,与单纯t细胞杀伤活性相比较,p53-t的肿瘤杀伤活性提高了12.5个百分点。
附:参考文献
1.meek,d.w.andc.w.anderson,posttranslationalmodificationofp53:cooperativeintegratorsoffunction.coldspringharbperspectbiol,2009.1(6):p.a000950.
2.appella,e.andc.w.anderson,post-translationalmodificationsandactivationofp53bygenotoxicstresses.eurjbiochem,2001.268(10):p.2764-72.
3.isobe,m.,etal.,localizationofgeneforhumanp53tumourantigentoband17p13.nature,1986.320(6057):p.84-5.
4.cho,y.,etal.,crystalstructureofap53tumorsuppressor-dnacomplex:understandingtumorigenicmutations.science,1994.265(5170):p.346-55.
5.hollstein,m.,etal.,p53mutationsinhumancancers.science,1991.253(5015):p.49-53.
6.carondefromentel,c.andt.soussi,tp53tumorsuppressorgene:amodelforinvestigatinghumanmutagenesis.geneschromosomescancer,1992.4(1):p.1-15.
7.fields,s.ands.k.jang,presenceofapotenttranscriptionactivatingsequenceinthep53protein.science,1990.249(4972):p.1046-9.
8.iwabuchi,k.,etal.,useofthetwo-hybridsystemtoidentifythedomainofp53involvedinoligomerization.oncogene,1993.8(6):p.1693-6.
9.jenkins,j.r.,k.rudge,andg.a.currie,cellularimmortalizationbyacdnacloneencodingthetransformation-associatedphosphoproteinp53.nature,1984.312(5995):p.651-4.
10.olivier,m.,m.hollstein,andp.hainaut,tp53mutationsinhumancancers:origins,consequences,andclinicaluse.coldspringharbperspectbiol,2010.2(1):p.a001008.
11.hainaut,p.andm.hollstein,p53andhumancancer:thefirsttenthousandmutations.advcancerres,2000.77:p.81-137.
12.kropveld,a.,etal.,sequencinganalysisofrnaanddnaofexons1through11showsp53genealterationstobepresentinalmost100%ofheadandnecksquamouscellcancers.labinvest,1999.79(3):p.347-53.
13.brown,c.j.,etal.,awakeningguardianangels:druggingthep53pathway.natrevcancer,2009.9(12):p.862-73.
14.inoue,k.,etal.,overexpressionofp53proteininhumantumors.medmolmorphol,2012.45(3):p.115-23.
15.iggo,r.,etal.,increasedexpressionofmutantformsofp53oncogeneinprimarylungcancer.lancet,1990.335(8691):p.675-9.
16.linzer,d.i.anda.j.levine,characterizationofa54kdaltoncellularsv40tumorantigenpresentinsv40-transformedcellsanduninfectedembryonalcarcinomacells.cell,1979.17(1):p.43-52.
17.lane,d.p.andl.v.crawford,tantigenisboundtoahostproteininsv40-transformedcells.nature,1979.278(5701):p.261-3.
18.flaman,j.m.,etal.,thehumantumoursuppressorgenep53isalternativelysplicedinnormalcells.oncogene,1996.12(4):p.813-8.
19.cutilli,t.,etal.,p53asaprognosticmarkerassociatedwiththeriskofmortalityfororalsquamouscellcarcinoma.oncollett,2016.12(2):p.1046-1050.
20.drosten,m.andm.barbacid,rasandp53:anunsuspectedliaison.molcelloncol,2016.3(2):p.e996001.
21.kung,c.p.andm.e.murphy,theroleofthep53tumorsuppressorinmetabolismanddiabetes.jendocrinol,2016.
22.lane,d.p.ands.benchimol,p53:oncogeneoranti-oncogene?genesdev,1990.4(1):p.1-8.
23.matlashewski,g.,etal.,isolationandcharacterizationofahumanp53cdnaclone:expressionofthehumanp53gene.emboj,1984.3(13):p.3257-62.
24.wolf,d.,etal.,isolationofafull-lengthmousecdnaclonecodingforanimmunologicallydistinctp53molecule.molcellbiol,1985.5(1):p.127-32.
25.fujita,k.,etal.,p53isoformsdelta133p53andp53betaareendogenousregulatorsofreplicativecellularsenescence.natcellbiol,2009.11(9):p.1135-42.
26.song,w.,etal.,expressionofp53isoformsinrenalcellcarcinoma.chinmedj(engl),2009.122(8):p.921-6.
27.hofstetter,g.,etal.,then-terminallytruncatedp53isoformdelta40p53influencesprognosisinmucinousovariancancer.intjgynecolcancer,2012.22(3):p.372-9.
28.bourdon,j.c.,etal.,p53isoformscanregulatep53transcriptionalactivity.genesdev,2005.19(18):p.2122-37.
29.nutthasirikul,n.,etal.,ratiodisruptionofthe133p53andtap53isoformequilibriumcorrelateswithpoorclinicaloutcomeinintrahepaticcholangiocarcinoma.intjoncol,2013.42(4):p.1181-8.
30.anensen,n.,etal.,adistinctp53proteinisoformsignaturereflectstheonsetofinductionchemotherapyforacutemyeloidleukemia.clincancerres,2006.12(13):p.3985-92.
31.oren,m.,p.tal,andv.rotter,targetingmutantp53forcancertherapy.aging(albanyny),2016.8(6):p.1159-60.
- 还没有人留言评论。精彩留言会获得点赞!
- 一种制备自噬小体型瘤苗的培养基添加剂及其制备方法与制造工艺
- N-萘烷基取代的含氮杂环衍生物、制备方法及其抗肿瘤用途与制造工艺
- 作为BET蛋白抑制剂的1H‑吡咯并[2,3‑c]吡啶‑7(6H)‑酮和吡唑并[3,4‑c]吡啶‑7(6H)‑酮的制造方法与工艺
- 一种人肿瘤抑制蛋白变体及其应用的制造方法与工艺
- 一种人肿瘤抑制蛋白变体及其应用的制造方法与工艺
- G蛋白信号调节因子6及其抑制剂在治疗脂肪肝和Ⅱ型糖尿病中的功能和应用的制造方法与工艺
- 蛋白酶体抑制剂在制备降低动物脂含量或脂储积能力产品中的应用的制造方法与工艺
- GINS2基因或蛋白的抑制剂在制备抗肿瘤药物中的应用的制造方法与工艺
- ZNF383蛋白在制备抑制p53蛋白活性的产品中的应用的制造方法与工艺
- 蛋白酶体抑制剂MLN9708的合成方法与制造工艺