一种抗DDX5的全人源单克隆抗体及其制备方法和应用与流程
本发明涉及一种全人源单克隆抗体及其制备方法和应用,特别涉及一种抗DDX5的全人源单克隆抗体及其制备方法和在检测DDX5表达以及预防或治疗DDX5相关的疾病中的用途,本发明属于医药技术领域。
背景技术:
DDX5(DEAD-Box Helicase 5)隶属于DEAD box家族,又称为p68,是一种ATP依赖的RNA解旋酶,最初被认为是一种参与细胞生长和分化的核蛋白。RNA解旋酶(RNA helicases)是一个包含了与RNA代谢(翻译起始、核糖体形成、前mRNA拼接和mRNA降解)许多方面有关的蛋白质家族。RNA解旋酶是由水解ATP供给能量来解开RNA的酶。它们常常依赖于单链的存在,并能识别复制单链结构。一般在RNA复制过程中起到催化双链RNA解旋的作用。DDX5同时具有依赖于RNA的ATPase活性,它与SV40大T抗原有交叉免疫反应。
DDX5在RNA代谢的各个方面发挥重要作用,参与mRNA前体加工、RNA反转、RNA转运、核糖体的合成和翻译以及RNA降解等过程。同时DDX5转录后的磷酸化也参与了肿瘤的发生和细胞的增殖和转化。例如,蛋白的SUMO(sumoylation)修饰可以直接影响DDX5的蛋白稳定性及转录功能。DDX5与HDAC1结合能够抑制转录,这种转录抑制的过程能够被SUMO修饰所调节。DDX5还能与Runx2,p53,Smad3,CBP以及p300相互作用。
已有研究表明,DDX5在乳腺癌、前列腺癌、结肠癌、急性髓性白血病等多种肿瘤细胞中高表达,抑制DDX5的表达能够抑制这些种类肿瘤细胞的增殖。DDX5在调控丙型肝炎病毒复制过程中亦发挥一定作用,说明DDX5的监测和调控在实现肿瘤和感染性疾病的精准诊断和治疗中具有重要价值。
目前全人源单克隆抗体是治疗性抗体发展的主要方向,本发明通过人源杂交瘤技术获得了全人源抗体克隆库,为抗体的制备筛选提供了良好的技术平台,筛选出来的抗体为人体内自然产生的抗体,避免了人源化过程引入的人为加工因素,更符合人体内的自然生理过程,具有广泛的应用前景。
技术实现要素:
本发明的目的在于提供一种抗DDX5的全人源单克隆抗体及其制备方法和应用。
为了达到上述目的,本发明采用了以下技术手段:
(1)抗DDX5单克隆抗体的初步筛选,通过ELISA技术对全人源杂交瘤抗体库中的抗体进行筛选,获得了一种亲和力较高的抗体。
(2)扩大培养筛选到的人源杂交瘤细胞株,亲和层析纯化单克隆抗体,通过免疫印迹技术,检测筛选到的抗体与DDX5纯化蛋白之间的亲和力;
(3)对筛选到的杂交瘤细胞进行抗体全基因测序,构建抗体DNA重组表达质粒。
(4)抗体DNA重组表达质粒转染宿主细胞,表达并纯化单克隆抗体,进行亲和力鉴定和比较。
(5)将上述(2)、(4)中获得的单克隆抗体与肿瘤细胞作用进行生物学功能鉴定和比较。
在上述研究的基础上,本发明筛选得到一株能够特异性结合DDX5的全人源单克隆抗体,其由轻链和重链组成,所述轻链可变区的氨基酸序列为SEQ ID NO.6所示,所述重链可变区的氨基酸序列为SEQ ID NO.8所示。
其中,轻链1G3-L1的互补决定区CDR1、CDR2和CDR3分别用LCDR1、LCDR2和LCDR3表示;LCDR1序列为QSVSSN,LCDR2序列为GAS,LCDR3序列为QQYNNWPRT。其中,重链1G3-H1的互补决定区CDR1、CDR2和CDR3分别用HCDR1、HCDR2和HCDR3表示;HCDR1序列为GFTFSDYS,HCDR2序列为ITSSSGYT,HCDR3序列为ARVRSSWGPIDS。
在本发明中,优选的,所述的特异性结合DDX5的全人源单克隆抗体中,轻链的氨基酸序列如SEQ ID NO:3所示,所述重链的氨基酸序列如SEQ ID NO:4所示。
本领域技术人员将知晓的是,抗体中的保守性氨基酸替换并不会实质性地影响抗体的亲和力和结构,尤其是所述保守性替换发生在恒定区时。因此,若氨基酸的变化发生在恒定区且变化后并不影响抗体的生物学活性,那么该抗体也应在本发明的保护范围之内。
进一步的,本发明还提出了包含所述全人源单克隆抗体的单个重链和/或单个轻链的单域抗体、嵌合抗体、抗体融合蛋白抗体、抗体/抗体片段-因子融合蛋白或抗体/抗体片段-化学偶联物。以及
所述的全人源单克隆抗体的抗原结合片段,其中,所述抗原结合片段可选自Fab、Fab'、F(ab')2、scFv、Fv、dsFv、双抗体、Fd和Fd'片段。
再进一步的,本发明还提出了一种分离的DNA分子,其为编码所述的抗体或所述的抗原结合片段的核苷酸序列,优选的,所述的核苷酸序列包含SEQ ID NO:5和/或SEQ ID NO:6所示的核苷酸序列,更优选的,所述的核苷酸序列包含SEQ ID NO:1和/或SEQ ID NO:2所示的核苷酸序列。
含有所述的核苷酸序列或其组合的重组DNA表达载体以及含有所述的核苷酸序列或所述的重组DNA表达载体的宿主细胞也在本发明的保护范围之内,优选的,所述的宿主细胞选自原核细胞、酵母或哺乳动物细胞,更优选的,所述宿主细胞选自HEK293F细胞、CHO细胞或NSO细胞。
所述HEK293F细胞为人胚肾293F细胞(human embryonic kidney 293F cell);CHO细胞为中国仓鼠卵巢细胞(chinese hamster ovary cell);NSO细胞为小鼠NSO胸腺瘤细胞(mouse thymoma NSO cells)。
更进一步的,本发明还提出了所述的抗体、所述的抗原结合片段、所述的核苷酸序列、所述的载体以及所述的宿主细胞在制备用于预防或治疗与DDX5相关的肿瘤、感染性疾病或免疫系统疾病,其中,优选的,所述肿瘤包括但不限于胶质瘤、霍奇金淋巴瘤、滤泡性淋巴癌、黑色素瘤、急性或慢性白血病、肺癌、乳腺癌、肝癌、胃癌、胰腺癌、肾癌、前列腺癌、膀胱癌、卵巢癌、皮肤癌、鳞状细胞癌、骨癌、头颈癌、结直肠癌、实体瘤;所述感染性疾病包括不限于HIV病毒感染、A型,B型或C型肝炎病毒感染、疱疹病毒感染、流感病毒感染;所述免疫系统疾病包括不限于系统性红斑狼疮、类风湿关节炎、强直性脊柱炎、重症肌无力、多发性硬化症、自身免疫性溶血性贫血、自身免疫性肝炎、硬皮病、结节性多动脉炎、Wegener肉芽肿病。以及
所述的抗体、所述的抗原结合片段、所述的核苷酸序列、所述的载体以及所述的宿主细胞在制备DDX5检测试剂中的应用。
更进一步的,本发明还提出了制备所述特异性结合DDX5的全人源单克隆抗体的方法,包括如下步骤:
(1)提供表达载体,所述表达载体含有编码所述抗体或所述抗原结合片段的DNA分子,以及与所述DNA分子可操作相连的表达调控序列;
(2)用所述表达载体转化宿主细胞;
(3)在适合表达所述抗体的条件下,培养含有所述表达载体的宿主细胞;
(4)分离纯化获得所述的单克隆抗体;
其中,优选的,所述的表达载体为含有抗体分泌信号肽序列的真核表达载体pcDNA3.4,所述的宿主细胞为HEK293F细胞。
本发明提供的单克隆抗体可以通过消除、抑制DDX5活性来预防、诊断和治疗疾病,本发明提供的抗DDX5蛋白单克隆抗体是全人源的,与其它动物源性(如鼠源性)的抗DDX5抗体相比,因物种差异导致的免疫原性大大减少,且特异性好、亲和力高,如果用于临床将大大降低副作用。此外,由于本发明中的单克隆抗体可以和DDX5抗原特异性结合,在DDX5的诊断和检测方面也具有良好的应用前景。
附图说明
图1为pcDNA3.4质粒图谱;
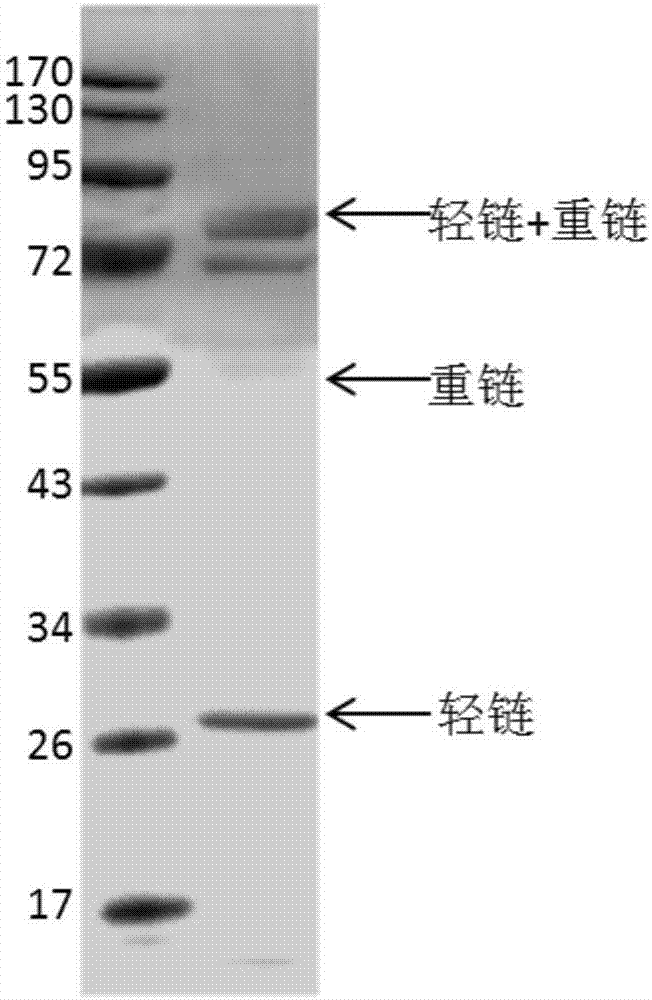
图2为杂交瘤细胞抗体表达蛋白电泳图;
图3为抗体DNA重组表达载体在宿主细胞中蛋白表达电泳图;
图4为单克隆抗体与DDX5在分子水平上的结合实验;
图5为单克隆抗体ELISA鉴定的亲和力;
图6为抗DDX5的单克隆抗体细胞内结合实验;
图7为抗DDX5的单克隆抗体对肿瘤细胞的抑制实验。
具体实施方式
本发明详细的实施方法参见实施例,实施例中所述的实验方法和试剂,若无特殊说明均为常规实验方法和试剂。以下实施例仅用于说明和解释本发明,而不是以任何方式限制本发明。
实施例1、抗DDX5单克隆抗体筛选
采用人源B细胞克隆技术建立全人源单克隆抗体库。以纯化的DDX5蛋白为抗原包被ELISA反应板,4℃包被过夜。再用4%的BSA/PBST封闭反应板,室温1小时。封闭后向反应板中加入抗体库杂交瘤细胞培养上清,室温反应1小时。PBST洗3次,每次5分钟。加入山羊抗人辣根过氧化物酶标记的二抗,室温1小时。PBST-PBS洗3次,每次5分钟。显色筛选阳性反应的杂交瘤细胞株,命名为1G3。
实施例2、抗DDX5的单克隆抗体纯化
实施例1得到的与DDX5蛋白反应阳性的杂交瘤细胞株1G3进行扩大培养,收集无血清细胞培养上清,蛋白电泳检测抗体表达,如图2。经过protein A亲和柱纯化,脱盐和浓缩,获得纯化的单克隆抗体。
实施例3、抗体DNA重组质粒构建与表达
将实施例1得到的与DDX5反应阳性的杂交瘤细胞1G3进行抗体全基因测序,获得抗体轻链和重链的基因序列:所述轻链(1G3-L1)的核苷酸序列为SEQ ID NO.1,氨基酸序列如SEQ ID NO.3所示,所述重链(1G3-H1)的核苷酸序列为SEQ ID NO.2,氨基酸序列如SEQ ID NO.4所示。
其中,轻链可变区核苷酸酸序列为SEQ ID NO.5,对应氨基酸序列为SEQ ID NO.6。轻链1G3-L1的互补决定区CDR1、CDR2和CDR3分别用LCDR1、LCDR2和LCDR3表示;LCDR1序列为QSVSSN,LCDR2序列为GAS,LCDR3序列为QQYNNWPRT。
其中,重链可变区核苷酸酸序列为SEQ ID NO.7,对应氨基酸序列为SEQ ID NO.8。重链1G3-H1的互补决定区CDR1、CDR2和CDR3分别用HCDR1、HCDR2和HCDR3表示;HCDR1序列为GFTFSDYS,HCDR2序列为ITSSSGYT,HCDR3序列为ARVRSSWGPIDS。
将上述轻链和重链基因分别克隆至装有抗体分泌信号肽序列的真核表达载体pcDNA3.4(图1)中。瞬时转染HEK293F细胞,进行全抗体表达,蛋白电泳检测抗体表达,如图3。使用protein A亲和柱纯化获得抗体蛋白。
实施例4、对筛选出的单克隆抗体与DDX5在分子水平上进行结合检测
DDX5纯化蛋白经10%SDS-PAGE电泳,转印到硝酸纤维素膜上,5%脱脂奶粉封闭,室温1小时。将实施例2纯化后的单克隆抗体用PBST稀释到1ng/μL,室温与膜孵育1小时。PBST洗3遍,每次5分钟。用anti-human Fc-HRP二抗孵育,室温1小时。PBST-PBS洗3遍。ECL显色,结果如图4。图4结果表明,实施例2纯化后的单克隆抗体能够与DDX5纯化蛋白结合。
实施例5、对筛选出的抗DDX5的单克隆抗体进行体外亲和力检测
ELISA检测实施例2纯化后的单克隆抗体与DDX5的亲和力,反应板包被纯化蛋白DDX5,4%BSA/PBST封闭4℃过夜,与不同浓度(1000ng/mL~0.001ng/mL)的单克隆抗体孵育,室温1小时。PBST洗3遍,每次5分钟。加入anti-human Fc-HRP二抗孵育,室温1小时。PBST-PBS洗3遍,每次5分钟。DAB显色,酶标仪读取OD450吸收值,结果如图5。图5结果表明,随着单克隆抗体浓度的下降,OD450吸收值也随之降低,说明纯化后的单克隆抗体能够与DDX5纯化蛋白结合。
实施例6、抗DDX5的单克隆抗体细胞内结合实验
常规培养人胶质瘤细胞,1×104cell/孔接种细胞培养皿,培养过夜,多聚甲醛固定,加入实施例2纯化后的抗DDX5的单克隆抗体,37℃反应1小时。PBST洗3次每次5分钟。加入Anti-human Fc-FITC二抗37℃反应1小时。PBST洗3次,每次5分钟。共聚焦显微镜观察实验结果。如图6。图6结果表明,纯化后的单克隆抗体能够与人胶质瘤细胞结合。
实施例7、抗DDX5的单克隆抗体对肿瘤细胞抑制实验
常规培养人胶质瘤细胞,1×104cell/孔接种细胞培养皿,培养过夜,加入实施例2纯化后的抗DDX5的单克隆抗体使其终浓度达到30μg/μL,培养不同时间取细胞,MTT法检测细胞活力,绘制细胞生长曲线。如图7。图7结果表明,随着孵育时间的延长,细胞活力下降,说明纯化后的单克隆抗体能够抑制人胶质瘤细胞的增殖及生长。
对于本领域的普通技术人员而言,具体实施例只是对本发明进行了示例性描述,显然本发明具体实现并不受上述方式的限制,只要采取了本发明的方法构思和技术方案进行的各种非实质性改进,或未经改进将本发明的构思和技术方案直接应用于其他场合的,均在本发明的保护范围之内。
序列表
<110> 哈尔滨医科大学
<120> 一种抗DDX5的全人源单克隆抗体及其制备方法和应用
<130> KLPI170019
<160> 8
<170> PatentIn 3.5
<210> 1
<211> 702
<212> DNA
<213> 1G3-L1
<400> 1
atggaagccc cagcgcagct tctcttcctc ctgctactct ggctcccaga taccactgga 60
gaaatagtga tgacgcagtc tccagccacc ctgtctgtgt ctccagggga aagagccacc 120
ctctcctgca gggccagtca gagtgttagc agcaacttag cctggtacca gcagaaacct 180
ggccaggctc ccaggctcct catctatggt gcatccacca gggccactgg tatcccagcc 240
aggttcagtg gcagtgggtc tgggacagag ttcactctca ccatcagcag cctgcagtct 300
gaagattttg cagtttatta ctgtcagcag tataataact ggcctcggac gttcggccaa 360
gggaccaagg tggaaatcaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 420
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 480
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 540
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 600
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 660
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 702
<210> 2
<211> 1404
<212> DNA
<213> 1G3-H1
<400> 2
atggagtttg ggctgagctg ggttttcctt gttgctatta taaaaggtgt ccagtgtcag 60
gtgcagctgg tggagtctgg gggagacttg gtcaagcctg gagggtccct gagactctcc 120
tgtgcagcct ctggattcac cttcagtgac tactccatga actgggtccg ccagactcca 180
gggaaggggc tggagtggct ttcatacatc actagtagta gtgggtacac aaattatgta 240
gactctgtga agggccgatt caccatctcc agagacaacg ccaagaactc actgtatctg 300
caaatgaata gcctgagagc cgaggacacg gctgtctact actgtgcgag agtgaggagt 360
agctggggac ctattgactc ctggggccag ggaaccctgg tcaccgtctc ctcagcctcc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gagggtctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg taaa 1404
<210> 3
<211> 234
<212> PRT
<213> 1G3-L1
<400> 3
1 MET Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu
16 Pro Asp Thr Thr Gly Glu Ile Val MET Thr Gln Ser Pro Ala Thr
31 Leu Ser Val Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
46 Ser Gln Ser Val Ser Ser Asn Leu Ala Trp Tyr Gln Gln Lys Pro
61 Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Thr Arg Ala
76 Thr Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu
91 Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser Glu Asp Phe Ala Val
106 Tyr Tyr Cys Gln Gln Tyr Asn Asn Trp Pro Arg Thr Phe Gly Gln
121 Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val
136 Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
151 Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
166 Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
181 Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
196 Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
211 Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
226 Thr Lys Ser Phe Asn Arg Gly Glu Cys
<210> 4
<211> 468
<212> PRT
<213> 1G3-H1
<400> 4
1 MET Glu Phe Gly Leu Ser Trp Val Phe Leu Val Ala Ile Ile Lys
16 Gly Val Gln Cys Gln Val Gln Leu Val Glu Ser Gly Gly Asp Leu
31 Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
46 Phe Thr Phe Ser Asp Tyr Ser MET Asn Trp Val Arg Gln Thr Pro
61 Gly Lys Gly Leu Glu Trp Leu Ser Tyr Ile Thr Ser Ser Ser Gly
76 Tyr Thr Asn Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
91 Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln MET Asn Ser Leu
106 Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Val Arg Ser
121 Ser Trp Gly Pro Ile Asp Ser Trp Gly Gln Gly Thr Leu Val Thr
136 Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
151 Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
166 Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
181 Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
196 Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
211 Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
226 Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
241 Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
256 Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
271 Thr Leu MET Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
286 Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
301 Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
316 Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
331 His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
346 Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
361 Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
376 Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
391 Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
406 Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
421 Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
436 Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val MET His
451 Glu Gly Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
466 Pro Gly Lys
<210> 5
<211> 291
<212> DNA
<213> 1G3-L1可变区
<400> 5
gaaatagtga tgacgcagtc tccagccacc ctgtctgtgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agcaacttag cctggtacca gcagaaacct 120
ggccaggctc ccaggctcct catctatggt gcatccacca gggccactgg tatcccagcc 180
aggttcagtg gcagtgggtc tgggacagag ttcactctca ccatcagcag cctgcagtct 240
gaagattttg cagtttatta ctgtcagcag tataataact ggcctcggac g 291
<210> 6
<211> 97
<212> PRT
<213> 1G3-L1可变区
<400> 6
1 Glu Ile Val MET Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro
16 Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser
31 Ser Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg
46 Leu Leu Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala
61 Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile
76 Ser Ser Leu Gln Ser Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
91 Tyr Asn Asn Trp Pro Arg Thr
<210> 7
<211> 324
<212> DNA
<213> 1G3-H1可变区
<400> 7
caggtgcagc tggtggagtc tgggggagac ttggtcaagc ctggagggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt gactactcca tgaactgggt ccgccagact 120
ccagggaagg ggctggagtg gctttcatac atcactagta gtagtgggta cacaaattat 180
gtagactctg tgaagggccg attcaccatc tccagagaca acgccaagaa ctcactgtat 240
ctgcaaatga atagcctgag agccgaggac acggctgtct actactgtgc gagagtgagg 300
agtagctggg gacctattga ctcc 324
<210> 8
<211> 108
<212> PRT
<213> 1G3-H1可变区
<400> 8
1 Gln Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly
16 Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
31 Asp Tyr Ser MET Asn Trp Val Arg Gln Thr Pro Gly Lys Gly Leu
46 Glu Trp Leu Ser Tyr Ile Thr Ser Ser Ser Gly Tyr Thr Asn Tyr
61 Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
76 Lys Asn Ser Leu Tyr Leu Gln MET Asn Ser Leu Arg Ala Glu Asp
91 Thr Ala Val Tyr Tyr Cys Ala Arg Val Arg Ser Ser Trp Gly Pro
106 Ile Asp Ser
- 还没有人留言评论。精彩留言会获得点赞!