糖基转移酶UGT71C3在催化合成水杨酸甲酯糖苷中的应用的制作方法
本发明涉及一种糖基转移酶的应用,尤其涉及一种来源于拟南芥的糖基转移酶ugt71c3在催化合成水杨酸甲酯糖苷中的应用;属于生物技术领域。
背景技术:
水杨酸甲酯糖苷类化合物是水杨酸甲酯的糖基化修饰衍生物,具有解热、镇痛、消炎等药理学作用。虽然这类化合物与水杨酸有类似作用,但有研究初步显示水杨酸甲酯糖苷可能比水杨酸本身的作用效果更强,副作用更小,有希望被开发成具有竞争力的解热、镇痛、消炎类新药。因此,近几年来,对水杨酸甲酯糖苷的药理作用的研究越来越受到重视。例如,对水杨酸甲酯糖苷抗大鼠急性胸膜炎的作用研究,结果显示水杨酸甲酯糖苷可明显降低胸膜炎大鼠胸腔炎性渗出液的体积以及渗出液中的白细胞数量和蛋白质含量,同时对于胸腔炎性渗出液中的no、tnf-α、il-1β和pge2的含量均有不同程度的抑制,能不同程度地降低血浆中mda含量,升高sod活力,表明了水杨酸甲酯糖苷具有抗大鼠急性胸膜炎作用(黄超等,中国药理学通报,2013)。另外,有研究表明水杨酸甲酯糖苷对脂多糖诱导的小神经胶质细胞炎性反应也有明显的抑制作用(马晓玮等,全国老年性痴呆与相关疾病学术会议论文集2013)。令人兴奋的是,水杨酸甲酯糖苷还有可能被用于缓解老年痴呆症。例如,在app/ps1转基因老年痴呆小鼠6月龄后给予饲喂水杨酸甲酯糖苷(150和300mg.kg-1.d-1),检测学习记忆能力、大脑皮质神经元超微结构退行性变化和小胶质细胞活化。结果发现水杨酸甲酯糖苷能够显著提高小鼠学习记忆能力,减缓神经退行性病变,保护皮质神经元超微结构。在分子机制方面,水杨酸甲酯糖苷抑制可显著抑制炎症因子tnf-α和il-6的基因表达和分泌,可减少细胞上清液中no的生成,可抑制inos蛋白和基因的表达,可显著抑制cox-1和cox-2蛋白和基因的表达,并减少pge2的基因表达和分泌。提示msl对活化bv-2具有显著的抗炎作用。同时,水杨酸甲酯糖苷通过抑制ikk激酶磷酸化、iκbα蛋白泛素化降解、以及p65蛋白核转位而抑制nf-κb信号通路的活化,发挥抗炎作用。可见水杨酸甲酯糖苷在app/ps1转基因老年痴呆小鼠模型中,可以通过抑制炎性因子的表达和抑制nf-κb信号通路的活化发挥抗炎作用,从而提高认知能力,保护神经细胞,对延缓ad病理损伤和改善学习记忆能力发挥了潜在作用(张天泰等,第十六届全国神经精神药理学学术会议论文集2014)。
近年来有关植物系统获得性免疫(sar)的研究发现,水杨酸甲酯在植物的sar反应中发挥了重要作用,其中park等(science,2007)人研究证明,如果在系统组织部位缺少mesa将导致植物的sar减弱甚至消失,随后liu等(plantphysiology,2011)也证明了这一观点。mesa作为植物体内的次级代谢物,在代谢途径中位于sa的下游,有甲基转移酶催化sa的甲基化产生mesa,同时又可以由水解酶水解变为sa,在植物体中的sa与mesa存在动态平衡。但是,在植物体内mesa是否存在糖基化修饰现象?催化mesa糖基化的酶是什么?mesa的糖基化修饰对于获得性免疫有什么作用?一直还缺乏了解。要回答这些问题,必须鉴定mesa的糖基转移酶和了解mesag在植物体内的生理学作用。
无论是用于医学研究还是植物学研究,要想获得水杨酸甲酯糖苷,目前主要还是依靠植物组织提取方法和化学合成方法。从植物体内提取,不仅量少而且很难分离到比较纯的目的糖苷;而化学合成的方法,费时费力,成本较高,污染较重,不是一个可持续发展的方法。相比之下,应用生物酶法合成水杨酸甲酯糖苷则具有高效高产、节约成本、环境友好等优点。
申请人通过大规模筛选拟南芥的100多个糖基转移酶,发现了一个能够催化合成水杨酸甲酯糖苷的糖基转移酶及其编码基因。该酶可以用于酶法合成水杨酸甲酯糖苷,为工业制备水杨酸甲酯糖苷提供了有力工具。经检索发现:到目前为止,有关利用糖基转移酶合成水杨酸甲酯糖苷的应用还未有报道。
技术实现要素:
针对现有技术的不足,本发明的目的是提供一种糖基转移酶ugt71c3在催化合成水杨酸甲酯糖苷中的应用。
本发明所述糖基转移酶ugt71c3在催化合成水杨酸甲酯糖苷中的应用。
其中:所述糖基转移酶ugt71c3,来自于拟南芥,由糖基转移酶基因ugt71c3编码其氨基酸序列,该基因的genbank号是nm_100600。
上述糖基转移酶基因ugt71c3是通过rt-pcr方法从拟南芥中克隆获得。上述糖基转移酶ugt71c3是通过糖基转移酶基因ugt71c3在大肠杆菌中表达和纯化后获得。
其中:所述糖基转移酶ugt71c3合成水杨酸甲酯糖苷的应用是以udp-葡萄糖为糖供体,利用糖基转移酶ugt71c3离体催化或在植物体内催化水杨酸甲酯(methylsalicylate)合成水杨酸甲酯糖苷。
具体的,将糖基转移酶ugt71c3与水杨酸甲酯共放置于反应管中,按实施例2中所给出的条件进行酶促反应,即可获得催化产物水杨酸甲酯糖苷(其与已经报道的其它糖基转移酶不同的是,本发明所述糖基转移酶ugt71c3能催化水杨酸甲酯合成糖苷,其它酶不能)。或者,将糖基转移酶基因ugt71c3在拟南芥超表达,制备成转基因植物,外源施加水杨酸甲酯,在转基因植物体内能够合成比对照植物更高含量的水杨酸甲酯糖苷(进一步证明了糖基转移酶ugt71c3可被用来催化合成水杨酸甲酯糖苷)。
本发明体现出的有益效果是:本专利在国内外首次公开了拟南芥糖基转移酶ugt71c3在以葡萄糖糖为糖供体,催化水杨酸甲酯合成水杨酸甲酯糖苷中的应用。为通过体外酶催化的方法合成水杨酸甲酯糖苷提供了条件,实现了应用生物酶法合成水杨酸甲酯糖苷(见附图),克服了用提取方法或化学合成方法获得水杨酸甲酯糖苷物效率低下,费时费力的不足。本发明实施后将为水杨酸甲酯糖苷工业化合成带来巨大经济效益。
附图说明
图1.纯化的融合蛋白gst-ugt71c3用sds-page检测。
其中,m为蛋白质分子量标记;序号1泳道为gst标签蛋白,作为对照;序号2泳道为纯化的gst-ugt71c3融合蛋白。
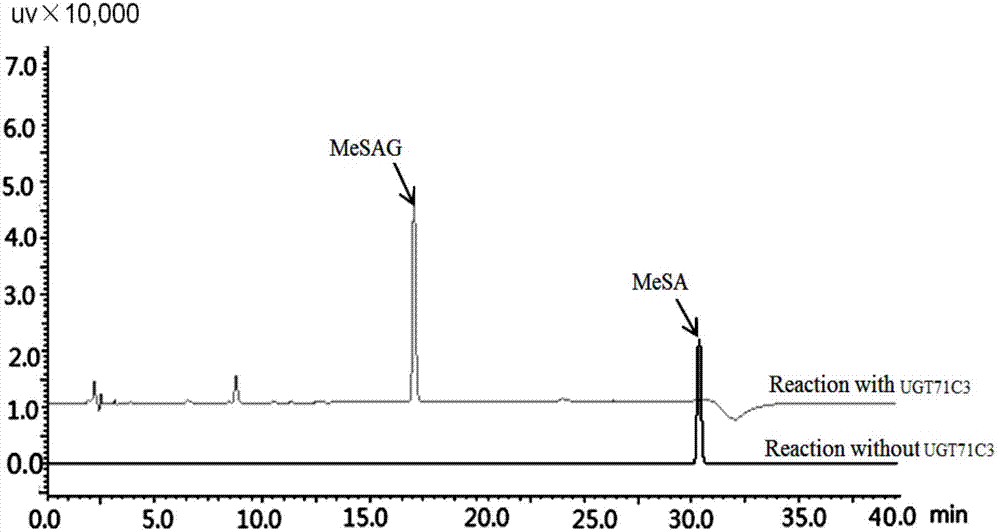
图2.纯化后的融合蛋白gst-ugt71c3催化水杨酸甲酯形成糖苷,催化产物用高效液相色谱(hplc)分析。
其中“reactionwithoutugt71c3”为对照组,仅用纯化的gst标签蛋白与水杨酸甲酯(图中mesa)和udp-葡萄糖反应,无糖苷合成;“reactionwithugt71c3”为实验组,用纯化的gst-ugt71c3融合蛋白与水杨酸甲酯和udp-葡萄糖反应,有糖苷合成(图中mesag)。
图3.催化合成的水杨酸甲酯糖苷用质谱(lc-ms)鉴定。
图中表示在正离子模式检测下,合成产物的主要的离子峰符合水杨酸甲酯糖苷的离子峰特征,证明水杨酸甲酯在ugt71c3酶的催化下形成了糖苷。
图4.通过转基因植物证明糖基转移酶ugt71c3能够催化水杨酸甲酯合成水杨酸甲酯糖苷。
图中wt为检测对照植物的水杨酸甲酯糖苷,oe6和oe10为检测糖基转移酶ugt71c3超表达转基因植物的水杨酸甲酯糖苷。给植物外源施加水杨酸甲酯以后一定时间,提取植物体内的糖苷物并通过高效液相色谱分析。发现在控制上样量均一的情况下(通过内标internalcontrol可反映),超表达ugt71c3的转基因植物比对照植物合成了更多的水杨酸甲酯糖苷(mesag)。证明糖基转移酶ugt71c3能够催化水杨酸甲酯合成水杨酸甲酯糖苷。
具体实施方式
实施例1拟南芥糖基转移酶基因ugt71c3的克隆、原核表达与酶蛋白纯化
通过筛选拟南芥的100多个糖基转移酶基因,发现ugt71c3编码的糖基转移酶能够催化水杨酸甲酯合成水杨酸甲酯糖苷。下面叙述该糖基转移酶的制作过程。
1.拟南芥糖基转移酶基因ugt71c3的克隆
通过公开的拟南芥专用数据库网站http://www.arabidopsis.org获得ugt71c3基因的cdna序列。根据cdna序列设计引物,正向引物为ugt71c3-f:5’-ggatccatgaaagcagaagcagagatcatc-3’,反向引物为ugt71c3-r:5’-gagctcctaaacatctccgccgatcaac-3’。为便于后续克隆,正向引物5’端添加bamhi酶切位点,反向引物5’端添加saci酶切位点。利用trizol试剂盒提取拟南芥rna,rt-pcr方法扩增ugt71c3基因的全长cdna序列。将cdna克隆的过程是先经过bamhi和sacⅰ酶切,之后连入相应酶切的pbluescriptiisk(+)载体中,构建成测序中间载体,称为pk-ugt71c3,然后用载体进行基因全长pcr扩增和bamhi和sacⅰ酶切验证,最后进行序列测定,验证克隆序列的正确性。
2.ugt71c3的序列信息与特性
对克隆的糖基转移酶基因ugt71c3测序后发现,该基因的编码区cdna为1431bp,编码476个氨基酸的蛋白,c端具有44个氨基酸的pspg保守序列盒,为植物次生代谢物糖基转移酶所共同具有的保守序列。
3.拟南芥糖基转移酶基因ugt71c3的原核表达载体构建
在公开通用的原核表达载体pgex-2t的基础上,按照《分子克隆》(sambrook等编著)所示的方法,对该载体进行改造,使其多克隆位点包含bamhi和saci。用bamhi和saci双酶切该表达载体,回收载体片段备用。同时用bamhi和saci双酶切前述载体pk-ugt71c3,回收得到目的基因片段,并将其连入上述原核表达载体中,得到拟南芥糖基转移酶基因ugt71c3的原核表达载体pgex-ugt71c3。
4.转化大肠杆菌、诱导蛋白表达及酶蛋白的纯化
将上述原核表达载体pgex-ugt71c3转化大肠杆菌菌株xl1-blue,pcr和酶切验证正确后,保存菌种,此菌种用于融合蛋白(gst-ugt71c3)的表达纯化。
在添加了氨苄青霉素(100mg/l)的2×yt液体培养基中过夜培养上述菌种,以活化该菌种。然后将过夜培养物按照1:100(过夜培养物:新的2×yt液体培养基)的比例扩大至75ml培养液中,37℃培养至od600=0.8。然后加入iptg至0.2mm,转到20℃震荡培养24h以诱导目的蛋白表达。到时间后通过离心收集菌体。
菌体加入2mlblastbuffer(0.5mmedta,750mm蔗糖,200mmtris-hcl,ph=8.0)重悬,立即加入14ml稀释的(1/2×)blastbuffer(事先添加2mg溶菌酶),混匀后于冰上放置30min,离心收集菌体,用pbs(内含0.2mm的pmsf)重悬,冰上放置30min后10000rpm离心20min,取上清。在上清中加入100μl的谷胱甘肽琼脂糖珠子,按照pgex蛋白纯化手册操作,最后用elutionbuffer(50mmtris-hcl,10mm还原型谷胱甘肽,ph=8.0)洗脱目的蛋白。目的蛋白的纯度用sds-page方法检测(附图1),蛋白浓度按照bradford试剂盒说明书中的方法进行测定(用bsa作为蛋白标准)。
实施例2糖基转移酶ugt71c3催化水杨酸甲酯生成水杨酸甲酯糖苷
1.水杨酸甲酯的酶催化反应
用纯化的糖基转移酶ugt71c3融合蛋白(gst-ugt71c3,能够反映ugt71c3的催化活性,属于常用的做法)进行体外的酶催化反应,以水杨酸甲酯作为底物,以udp-glucose作为糖供体。
酶促反应体系如下:
将以上混合好的反应体系放于30℃恒温水浴锅中,反应3小时。然后加入20μl浓度为240mg/ml的三氯乙酸终止反应。将反应管用液氮速冻后存放在-20℃,直到hplc分析。
2.酶催产物的hplc鉴定
用高效液相色谱(hplc)分析以上的反应混合体系,所用的仪器为岛津lc-20at,柱子为安捷伦公司的c18反相柱(ultimate,4.6×150mm,5μm)。a相为有机相(甲醇,含0.1%磷酸),b相为水相(含0.1%磷酸),设定色谱程序:最初(0min)b相为90%;20min达到b相60%;20.01min达到b相80%,维持至40min结束。hplc分析显示,与对照(反应体系中只含有纯化的gst标签蛋白)相比,只有ugt71c3融合蛋白能够催化水杨酸甲酯形成一种催化产物,推测是相应的水杨酸甲酯糖苷(附图2)。
3.酶催产物的质谱鉴定
对酶催化产物进行进一步的lc-ms鉴定,本试验采用的质谱色谱联用仪是surveyormsq(美国),检测方式为正离子模式检测,色谱程序同上述hplc程序。
结果证明酶促反应生成的产物确实为水杨酸甲酯糖苷(见附图3)。由此得出结论:糖基转移酶ugt71c3可用于催化合成水杨酸甲酯糖苷。
实施例3通过转基因植物证明糖基转移酶ugt71c3能够催化合成水杨酸甲酯糖苷
1.植物表达载体构建和转基因植物获得
1.1植物表达载体的构建
将测序正确的质粒pk-ugt71c3和植物表达载体pbi121用相同的酶(bamhi和saci)切开,分别回收基因片段和载体,进行连接,方法同原核表达载体的构建。
1.2植物表达载体转化农杆菌感受态
(1)取重组质粒5ul,加入到50ul的农杆菌gv3101感受态中,冰浴30min.
(2)液氮速冻1min,立即置于37℃的水浴锅中温育3min。
(3)冰浴3min后加入800ul的lb培养基,置于28℃的摇床中180rpm,5h。
(4)涂板,将复苏的农杆菌涂布到含有卡那霉素50mg/l和利福平50mg/l的平板中,置于28℃温箱中培养2天。
(5)采用pcr方法,验证表达载体已经转入农杆菌。
1.3含表达载体的农杆菌转化拟南芥
本试验采用的是浸花法转化拟南芥
(1)将鉴定的阳性单克隆置于28℃的摇床中扩大培养。
(2)收集菌体,4500rpm离心10min。
(3)弃上清,用5ml的5%蔗糖溶液(内含万分之二的silwet177)重悬菌体。
(4)测od600,在0.8~1.2即可用于浸染。
(5)用200ul的移液枪将重悬的菌体蘸到拟南芥未开的花序上,每隔30min一次,共浸染3次。
(6)用黑色塑料袋将浸染过的拟南芥罩住,过夜,以提供一个湿润的环境。
(7)以后的每周都要浸染,连续3周。
1.4转基因植株的获得
待浸染的拟南芥成熟后,收获种子,放置于37℃的温箱中使种子烘干,以利于贮存。
(1)种子的灭菌,将烘干的种子在超净工作台中进行灭菌,75%乙醇2min,1‰升汞3min,无菌ddh2o洗3遍。
(2)春化处理,将灭过菌的种子置于4℃冰箱中黑暗处理3天。
(3)将春化过的种子铺种到含有50mg/l卡那霉素的ms培养基上,培养14天。
(4)14天后将生长正常的拟南芥移到土中,待其长大后每一颗都单独收种子,在按照上述步骤,筛选绿苗:黄苗=3:1的株系,将绿苗移到土中,收获种子后,再用抗生素平板筛选纯合体。
(5)提取纯合体幼苗的rna通过rt-pcr或者qrt-pcr鉴定高表达株系。
1.5水杨酸甲酯糖苷的分析
(1)取14天大小的转基因纯合体拟南芥,置于含有100umol/l水杨酸甲酯(mesa)的ms培养液中,放在低速的摇床上处理48h,可设置不含mesa的培养液为对照组,48h后将植物取出,用吸水纸吸干水分,称重1g,液氮速冻。
(2)在液氮中将植物材料研碎,置于10ml的离心管中,立即加入90%的甲醇溶液,此时加入邻甲氧基苯甲酸作为内标。
(3)1700rpm离心十分钟,留上清,将上清转移至新的离心管中。
(4)过滤,过滤其中含有的杂质,
(5)将过滤所得的液体在真空泵中蒸干,用纯甲醇回溶,所得的溶液立即用于色谱分析。色谱分析条件同上述酶催产物的hplc鉴定。
(6)分析结果表明:超表达糖基转移酶ugt71c3的转基因植物比对照植物合成了更多的水杨酸甲酯糖苷。实验再次证明了糖基转移酶ugt71c3能够催化水杨酸甲酯合成水杨酸甲酯糖苷。
- 还没有人留言评论。精彩留言会获得点赞!
- 用于检测α-甲酰基-CoA消旋酶(AMACR)和前列腺癌的系统和方法
- 与赖氨酰氧化酶样蛋白2(loxl2)相关的治疗和诊断方法
- 包含3-甲酰基利福霉素sv和3-甲酰基利福霉素s的3-(4-肉桂基-1-哌嗪基)氨基衍生物的 ...的制作方法
- 用于检测赖氨酰氧化酶样蛋白-2(loxl2)和乳癌的系统和方法
- 一种用固定化脂质酰基转移酶对大豆毛油脱胶的方法
- 作为组织转谷氨酰胺酶的抑制剂的吡啶酮衍生物的制作方法
- 用于转移在具有空位团的基片中形成的薄层的改进方法
- 一种用于人血清谷氨酰基转移酶测定的缓冲液的制作方法
- 使用甘氨酰-tRNA合成酶和钙粘蛋白筛选用于预防或治疗癌症的试剂的方法
- 一种精浆γ-L-谷氨酰基转肽酶活性定量检测试剂盒及应用的制作方法