调控石榴花胚珠发育的PgAG基因及其用途的制作方法
本发明涉及一种从石榴中克隆得到调控石榴花胚珠发育的PgAG基因及其用途。
背景技术:
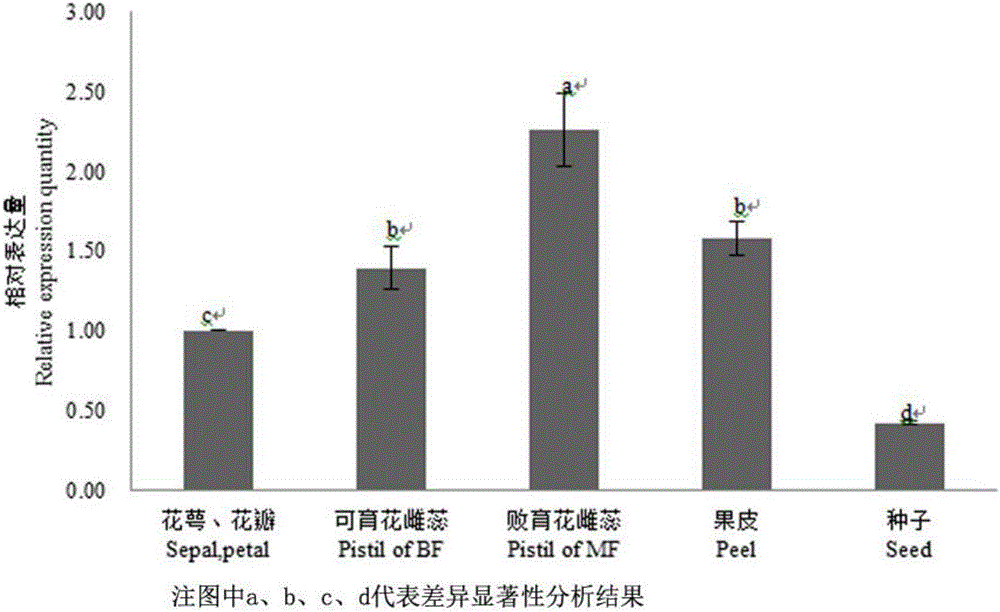
:石榴(PunicagranatumL.)起源于中亚地区,在世界各地广为栽培,具有很高的加工及保健价值。研究表明,石榴及其加工品对癌症、动脉粥样硬化、冠心病、高血压、高血脂、艾滋病、传染性疾病等具有重要的预防治疗作用。石榴花发育过程中存在严重雌蕊败育现象,而石榴花雌蕊败育是制约其果实产量的关键因素。控制植物雌蕊发育的途径主要分为两条:第一条为WUS-AG相关基因共同调控雌蕊发育;第二条为KNOXI类基因调控雌蕊发育。而第一条途径为主要调控途径。AG类基因在植物雌蕊发育过程中起重要作用。因此,克隆石榴中AG类同源基因及功能验证对石榴花雌蕊败育机理研究具有重要意义。Coen等1991年提出ABC模型,将AG类基因归类到C类基因,在花雄蕊及雌蕊发育过程中起着关键作用,ag突变体表现为雄蕊花瓣化,雌蕊萼片化。随后的研究发现,在ABC模型中包含调控胚珠发育的D类基因,及与所有花器官形成均相关的E类基因。因此,最初的ABC模型被扩展为ABCDE模型。ABCDE模型中的基因大部分属于MADS-BOX基因家族,其编码的氨基酸序列具有典型的MADS与Keratin-like保守结构域。AG类亚家族基因是MADS-BOX基因家族成员,而ABCD模型中C类与D类基因共属于AG类亚家族基因,在雌蕊发育过程中起着重要的作用。古代基因组复制事件使AG类亚家族基因分为两支:AGAMOUS分支和AGL11分支,C类基因一般归于AGAMOUS分支,AGL11单独形成一支,调控胚珠发育。随后AGL11在拟南芥、番茄、葡萄等物种上研究均证实其在胚珠发育及种子形成过程中的重要性。石榴花雌蕊败育的主要原因为胚珠发育异常,但其分子机理及AGL11在其分子调控过程中所起作用尚不知。随着对模式植物拟南芥等花发育方面的研究,了解石榴开花机理可有效地指导石榴育种,加快育种进程。但是迄今为止尚未见有关从石榴上分离PgAG基因对花发育影响的相关报道。技术实现要素:本发明提供一个首次从石榴中获得的的PgAG基因,该基因具有调控石榴花胚珠发育的功能。为解决上述技术问题,本发明采用的技术方案为:调控石榴花胚珠发育的PgAG基因,其核苷酸序列如:SEDIDNO:1。本发明的另一个方面:所述PgAG基因为完整的编码231个氨基酸的ORF,具有MADS-box基因的结构,编码的蛋白含有57个氨基酸的MADS结构域、82个氨基酸的K结构域。本发明的另一个方面:所述PgAG基因编码的氨基酸序列如SEQIDNO:2。本发明的另一个方面:经实时荧光定量表达分析,所述PgAG基因在石榴的败育花雌蕊、果皮、可育花雌蕊、花萼花瓣和种子中都有表达,其中,在雌蕊中的表达量高于别的部位;依据石榴可育花与败育花花蕾纵径大小不同分三个时期:I(花蕾纵径5-8mm),II(花蕾纵径8.1-15mm),III(花蕾纵径15-25mm),II为败育花败育的关键时期,II时期时PgAG基因在可育花雌蕊中表达量显著性高于败育花雌蕊。本发明的PgAG基因在调节拟南芥开花中的应用,所述PgAG基因与载体pBI121正向相连后得到PBI121-PgAG重组表达载体,将所述PBI121-PgAG重组表达载体转化农杆菌菌株GV3101,将拟南芥植株的地上部位倒置于浸染液中,暗培养过夜,即可正常栽培管理,一周以后可再侵染一次,等待收获T0代转基因种子,T1代植株的花瓣变小,雄蕊变短,花柱柱头雌蕊粗壮,表面乳突状细胞变长,石榴PgAG基因对拟南芥雌蕊发育具有正调控作用,同时对花瓣和雄蕊发育具有抑制作用。附图说明为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。图1为石榴PgAG基因序列与其他物种同源基因蛋白系统进化树与保守结构域预测结果图;图2不同花器官部位石榴PgAG基因表达量差异结果图;图3不同发育时期可育花与败育花雌蕊PgAG基因表达量差异结果图,图4为基因CDS全长序列克隆的PgAG电泳分析图;图5为过量表达载体的构建的PgAG电泳分析图;图6为PCR验证转基因植株的电泳分析图;图7转基因拟南芥与野生型拟南芥表型对比图。具体实施方式下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有作出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。本发明的PgAG基因是从突尼斯软籽石榴中克隆提取得到的,其核苷酸序列如:SEDIDNO:1(序列1),其氨基酸序列如SEDIDNO:2(序列2)。一、总RNA提取及第一链cDNA合成总RNA提取采用改良的CTAB法,用ThermoScientificNanodrop1000微量紫外可见光分光光度计测量RNA浓度及OD260/280值,并利用琼脂糖电泳检测确定RNA的完整性。cDNA第一链合成参照TIANscriptRTKit操作说明书。提取的总RNA作如下分析:核酸开放阅读框序列分析由软件DNAMAN完成;通过NCBIBLASTP对其核苷酸序列进行相似性分析;利用NCBICDD对其保守序列进行预测分析;蛋白质一级结构分析利用瑞士生物信息学研究所提供的ProtParam软件,在线分析氨基酸残基数目和组成、蛋白质相对分子质量、分子式、理论等电点及亲水性等理化性质分析,结果如图1所示。经分析所述PgAG基因为完整的编码231个氨基酸的ORF,具有MADS-box基因的结构,编码的蛋白含有57个氨基酸的MADS结构域、82个氨基酸的K结构域,与桉树、拟南芥、甜橙等物种AGL11相似性高。二、实时荧光定量PCR验证利用CTAB法提取不同发育时期雌蕊及盛花期不同花部位的RNA,利用TIANscriptRTKit进行cDNA第一链合成用于实时荧光定量PCR。设计PgAG荧光定量表达引物。上游引物:5’-GCAGCTAGAGAACAGGCTCG–3’,下游引物:5’-AAATTGCGGGAAGCCAACAC-3’。以石榴Actin作为内参基因,其引物上游引物:5’-AGTCCTCTTCCAGCCATCTC-3’,下游引物:5’-CACTGAGCACAATGTTTCCA-3’。用SYBRGreenI法进行real-timePCR定量分析,反应体系为SYBRGreenReal-timePCRMasterMix(2X)10μL;PCRForwardPrimer(10μmol/L)0.5μL;PCRReversePrimer(10μmol/L)0.5μL;cDNA模板2μL;加灭菌蒸馏水补至20μL。所用仪器为Roche480。反应程序为95℃5min;95℃10s,60℃10s,72℃10s,45个循环。相对表达量的计算采用2-△△Ct法。用Spass软件对其表达量进行方差分析。具体结果见图2和图3。如图2和3所示,经实时荧光定量表达分析,所述PgAG基因在石榴的败育花雌蕊、果皮、可育花雌蕊、花萼花瓣和种子中都有表达,其中,在雌蕊中的表达量高于别的部位;依据石榴可育花与败育花花蕾纵径大小不同分三个时期:I(花蕾纵径5-8mm),II(花蕾纵径8.1-15mm),III(花蕾纵径15-25mm),II为败育花败育的关键时期,II时期时PgAG基因在可育花雌蕊中表达量显著性高于败育花雌蕊,表明石榴PgAG基因对石榴影响石榴花败育。三、基因CDS全长序列克隆以盛花期突尼斯软籽石榴败育花雌蕊为材料,CTAB法提取RNA,采用TA克隆方式,依据石榴基因组数据库中PgAG基因的参考序列使用Primer-Primer6.0设计克隆引物。上游引物:5’-ATGGGGAGAGGAAAGATCGA-3’,下游引物:5’-CTAGACTAGTATGCATAGAATT-3’。PCR扩增采用NEB的Phusion超保真DNA聚合酶进行扩增。反应体系:5×PhusionHF缓冲液,10μL;2.5mMdNTP,4μL;模板cDNA2μL;上下游引物(10mmol·L-1)各2.5μL;Phusion超保真酶0.5μL;补灭菌ddH2O至50μL。PCR反应程序为:98℃30s;98℃10s,58℃10s,72℃30s,38个循环;72℃10min;4℃保存。PCR产物用2%的琼脂糖进行跑胶检测,凝胶回收用TakaraDNA凝胶回收试剂盒进行回收。纯化后的产物用TaqDNA聚合酶加A尾巴。反应体系:TaqDNA0.5μL;10×buffer2.5μL;25mMMg2+1.5μL;2.5mMdNTP2μL;模板补至25μL,反应程序:72℃10min。将纯化后的产物连接到pMD19-T载体上,转化到大肠杆菌DH5α感受态中,进行蓝白斑筛选,挑取阳性克隆送生工生物工程(上海)有限公司进行测序,测序通用引物为M13。上游引物:5’-gagaacacgggggacTCTAGAATGGGGAGAGGAAAGATCG-3’;下游引物:5’-gagaacacgggggacTCTAGAATGGGGAGAGGAAAGATCG-3’。具体步骤参照PMD19-T操作步骤,操作方法如下:1、在微量离心管中配制下列DNA溶液,全量为5μl。试剂使用量pMD19-TVector*11μlControlInsert*21μldH2O3μl2、加入5μl(等量)的SolutionI。3、16℃反应30分钟。4、全量(10μl)加入至100μlJM109感受态细胞中(换成DH5α感受态细胞),冰中放置30分钟。5、42℃加热45秒钟后,再在冰中放置1分钟。6、加入890μlSOC培养基,37℃振荡培养60分钟。7、在含有X-Gal、IPTG、Amp的L-琼脂平板培养基上培养,形成单菌落。计数白色、蓝色菌落。8、挑选白色菌落,使用PCR法确认载体中插入片段的长度大小。蓝白斑筛选后挑取阳性克隆,跑胶检验得到一段约750bp长度的序列(图4)。由图4看出条带2中有微弱的杂带,因此回收条带3,送生工生物科技测序,结果得到CDS全长序列696bp。上传NCBI,登录号为:KX378137,命名为PgAG。四、过量表达载体的构建:选用PBI121载体(该载体由生工生物工程(上海)有限公司提供,为连有PgAG目的片段的载体)。上游引物:5’-gagaacacgggggacTCTAGAATGGGGAGAGGAAAGATCG-3’;下游引物:5’-gagaacacgggggacTCTAGAATGGGGAGAGGAAAGATCG-3’。限制性酶切位点选用BamHI/XbalI,其在引物中的位置用下划线标出。利用上述引物扩增、构建带有35S强启动子的PBI121-PgAG重组表达载体,具体步骤如下:1、利用上述引物对连有PgAG基因的PMD19-T进行PCR扩增,琼脂糖跑胶回收的到PgAG基因片段;2、利用限制性内切酶BamHI/XbalI对PBI121载体进行酶切;3、T4连接酶连接PgAG基因片段与PBI121载体过夜;4、将重组质粒转化到DH5α大肠杆菌中;5、将其涂布于含卡那霉素的LB培养基中进行阳性筛选,将阳性菌落于含卡那霉素抗性的LB液体培养基中培养;6、提取质粒,进行跑胶、测序检验。取阳性克隆回收质粒,利用XbalI和BamHI双酶切法构建PBI121-PgAG重组质粒,跑胶检验(图5)并经生工测序检查植物过表达载体中的PgAG基因阅读框均未发生突变。这说明植物表达载体构建成功。五、农杆菌介导法转基因1、农杆菌转化取100uL农杆菌GV3101感受态细胞,加1ug重组质粒,冰浴30min;液氮速冻1-2min,放置于37℃轻摇5-6min,然后加入800ul新鲜的YEP液体培养基中;28℃,180rpm震荡培养2-4h,4000rpm离心5min,弃上清液,在沉淀中加入200ul新鲜YEP液体培养基,将其混匀,涂布于含50mg/L的卡那霉素和利福平的YEP固体培养基上(另设一个仅含有利福平的YEP平板作对照)。28℃培养2-3d,观察结果。挑取阳性菌落与1.5ml离心管中加入800ul含有卡那霉素和利福平的YEP液体培养基,28℃,220rpm震荡培养1天,得到含有目的质粒的农杆菌GV3101菌液。2、制侵染液在含有卡那霉素和利福平的YEP液体培养基中培养含有目的质粒的农杆菌GV3101(5ml液体培养基加10ul上述农杆菌GV3101菌液),28℃,220rpm过夜培养;取2-3ml液体培养物于100ml含卡那霉素和利福平的YEP液体培养基,28℃,220rpm震荡培养到OD600=1.2-1.6(大约4h),4000rpm离心20min,弃上清液,收集沉淀菌体,将菌体重悬于侵染介质(1/2MS培养基)中,使OD600=0.8左右,加表面活化剂silweetl-77。3、浸染将拟南芥植株的地上部位倒置于浸染液中1-2min,暗培养过夜,于光照培养箱(L23℃,14h/D18℃,10h)中培养,观查其生长表型,一周以后可在侵染一次,等待收获T0代转基因种子。五、转基因拟南芥植株获得收获转基因拟南芥T0代种子,消毒后点种于含100mg/L卡那霉素的1/2MS培养基上筛选阳性植株。8天后观察发现,阳性植株生长势强,呈深绿色,根系旺盛,而非阳性植株黄化枯萎直至死亡。取在含卡那霉素的1/2MS培养基上生长势良好的阳性苗叶片提取DNA,利用克隆时所用引物进行PCR验证采用CTAB法提取DNA,利用TaqDNA聚合酶进行PCR扩增,引物选用克隆引物,上游引物:5’-ATGGGGAGAGGAAAGATCGA-3’,下游引物:5’-CTAGACTAGTATGCATAGAATT-3’。扩增反应程序为:95℃5min;95℃30s,55℃30s,72℃1min,35个循环;72℃10min,4℃保存。扩增产物用1.5%的琼脂糖凝胶电泳分离检测。具体结果见图6。由图6可知经验证的11株转基因苗中10株跑出条带证明为转基因植株。将10株转基因植株同野生型对照植株一起于光照培养箱(L23℃,14h/D18℃,10h)中培养,观察T1代植株开花表型。六、转基因拟南芥表型鉴定将T0代转基因拟南芥种子点种于含100mg/L卡那霉素的1/2MS固体培养基上,同时,点种野生型拟南芥种子于1/2MS固体培养基上做对照。观察转基因拟南芥植株表型,观察期开花时间,花形态。利用GUS染液染色观察石榴PgAG基因在转基因拟南芥花中的主要表达部位。具体结果见图7。由图7可知,T1代植株表现为花瓣变小,雄蕊变短,雌蕊粗壮,花柱柱头表面乳突状细胞变长。结果表明石榴PgAG基因对拟南芥雌蕊发育具有正调控作用,同时对花瓣和雄蕊发育具有抑制作用。以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。SEQUENCELISTING<110>中国农业科学院郑州果树研究所<120>调控石榴花胚珠发育的PgAG基因及其用途<130>2016<160>2<170>PatentInversion3.3<210>1<211>696<212>DNA<213>石榴<400>1atggggagaggaaagatcgagatcaagaggatcgagaacacgacgaaccgtcaggtcacg60ttttgcaagcgtaggaatggattgctgaagaaagcgtatgagctctcagtgctctgcgat120gcggaagttgccctcatcgtcttttccagcagaggccgcctctatgagtactcgaacaat180aacatcaagacaactatagagaggtataagaaggctagctcagacagtgcgaacacgact240tctgtcatagagatcaatgctcagtattatcaacaagaatcagccaagctgaggcagcag300atacagatgctgcagaactctaacaggcatttgatgggagattctttgagtgccctttct360gtgaaggaactgaagcagctcgagaacaggctcgagcgcggaatcaccagaattaggtcc420aagaagcatgaaatgctgctttgtgagattgagtacttgcagaaaaaggagattgagatg480gaaaatgaaagtgtgtaccttcgcaccaagatagctgagatagagaggatggagcaagca540aacatggttccgggccaggagatgaatgcgattcatgtgttggcttcccgcaatttcttc600ccccccaatatgttggagggtgggaattcctactctcatcctgataagaaactccatctc660gggtatgaatcaataattctatgcatactagtctag696<210>2<211>231<212>氨基酸<213>石榴<400>2MetGlyArgGlyLysIleGluIleLysArgIleGluAsnThrThrAsn151015ArgGlnValThrPheCysLysArgArgAsnGlyLeuLeuLysLysAla202530TyrGluLeuSerValLeuCysAspAlaGluValAlaLeuIleValPhe354045SerSerArgGlyArgLeuTyrGluTyrSerAsnAsnAsnIleLysThr505560ThrIleGluArgTyrLysLysAlaSerSerAspSerAlaAsnThrThr65707580SerValIleGluIleAsnAlaGlnTyrTyrGlnGlnGluSerAlaLys859095LeuArgGlnGlnIleGlnMetLeuGlnAsnSerAsnArgHisLeuMet100105110GlyAspSerLeuSerAlaLeuSerValLysGluLeuLysGlnLeuGlu115120125AsnArgLeuGluArgGlyIleThrArgIleArgSerLysLysHisGlu130135140MetLeuLeuCysGluIleGluTyrLeuGlnLysLysGluIleGluMet145150155160GluAsnGluSerValTyrLeuArgThrLysIleAlaGluIleGluArg165170175MetGluGlnAlaAsnMetValProGlyGlnGluMetAsnAlaIleHis180185190ValLeuAlaSerArgAsnPhePheProProAsnMetLeuGluGlyGly195200205AsnSerTyrSerHisProAspLysLysLeuHisLeuGlyTyrGluSer210215220IleIleLeuCysIleLeuVal225230当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 拟南芥SSCD1基因突变在调控植物体内茉莉酸合成中的应用的制作方法与工艺
- 一种基因调控网络的优化控制方法与流程
- 拟南芥WRKY家族转录因子AtWRKY44基因,蛋白编码序列及其应用的制作方法与工艺
- 针对免疫调控能力相关基因个体化配方食品及其制备方法与流程
- REVOLUTA基因在调控豆科植物小叶片数目和叶茎比中的应用的制作方法与工艺
- 玉米锌铁调控转运体ZmZIP3基因及其应用的制作方法与工艺
- 拟南芥基因SPOC1在调控植株开花期中的应用的制作方法与工艺
- 调控石榴花胚珠发育的PgAG基因及其用途的制作方法与工艺
- 光效调控基因HPE1及其应用的制作方法与工艺
- 菘蓝IiAP2/ERF049基因在调控木脂素类化合物合成中的应用的制作方法与工艺