花期调控基因和相关载体及其应用的制作方法
本技术领域涉及植物育种和遗传学,特别是涉及用于调控植物开花期和/或抽穗期的重组dna构建体和调控植物开花期和/或抽穗期的方法。
发明背景
植物的生长周期通常包括营养生长期和生殖生长期,从营养生长到生殖生长的转变受到多种开花信号的影响,而开花信号受到诸如基因型的遗传因子和诸如光周期和光强度的环境因子等多种因素的影响(dung等,theoreticalandappliedgenetics,97:714-720(1998))。
开花期或抽穗期是重要的农业性状,是植物分布和地域适应性的重要决定因素。大部分被子植物品种响应诸如日照长度和温度等环境刺激和包括发育时期在内的内部信号的诱导开花。
从遗传的角度来看,植物中存在两种表型变化,控制营养生长和成花生长。第一种遗传改变包括从营养到成花状态的转换,如果这种遗传改变不能正常发挥作用,那么将不会出现开花;第二个遗传事件是形成花。植物器官的顺序发育的观察表明存在一个遗传机制调控一系列基因依次被的开启和关闭。
两个远亲双子叶植物拟南芥和金鱼草属植物研究,鉴定了三类同源异型基因,单独或联合决定花器官特征(bowman等,development,112:1(1991);carpenter和coen,genesdevl.,4:1483(1990);schwarz-sommer等,science,250:931(1990)),其中的一些基因是转录因子,其保守dna结合结构域被指定为mads盒(schwarz-sommer等,supra)。
控制花分生组织特征的早期活性基因也被分析确定。在拟南芥和金鱼草属植物中,花分生组织源于花序分生组织。控制分生组织细胞发育成花的两个因子是已知的,拟南芥中为leafy基因的产物(weige等,cell69:843(1992))和apetala1基因的产物(mandel等,nature360:273(1992))。当其中任何一个基因被突变失活时,花或花序的结构特征发生改变(weigel,etal.,supra;irish和sussex,plantcell,2:741(1990))。在金鱼草属植物中,与拟南芥leafy基因同源的基因为floricaula(coen等,cell,63:1311(1990))和apetala1基因的同源基因squamosa(huijser等,emboj.,11:1239(1992))。后两个因子包含mads盒结构域。
有效的开花在植物中是很重要的,尤其是当预期的产品是花或由此产生的种子时。促进或延缓开花的起始对农民或种子生产者是有用的,理解影响开花的遗传机制为改变目标植物开花特征提供了方法。多种植物中开花对作物生产非常重要,基本上所有农作物都是种子长成的,谷物是重要的例子,水稻和玉米是温带气候区最重要农艺作物,小麦、大麦、燕麦和黑麦是温带气候区的重要农作物。重要的种子产品是含油种子芸苔、加拿大油菜、糖用甜菜、玉米、向日葵、大豆和高粱。控制开花时间在园艺中很重要,开花时间可以控制的园艺作物包括莴苣、菊苣和包括卷心菜、花椰菜和菜花的蔬菜芸苔,和康乃馨和天竺葵。
发明概述
一方面,本发明包括一个分离的调控植物开花时间的多核苷酸,其包括:(a)一种多核苷酸,其核苷酸序列与seqidno:2和5的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:3和6的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4或7的序列一致性至少为90%;或(d)(a)、(b)或(c)核苷酸序列的全长互补序列,其中在植物中,过量表达所述的多核苷酸促进营养生长向生殖生长的转变,降低所述多核苷酸的表达量延长营养生长向生殖生长的转变时间。核苷酸序列包括其包括seqidno:2、seqidno:3、seqidno:5或seqidno:6的核苷酸序列。多肽的氨基酸序列包括seqidno:4或seqidno:7。
另一方面,本发明包括重组dna构建体,其包含分离的多核苷酸和与之可操作连接的至少一个调控序列,其中所述多核苷酸包含(a)一种多核苷酸,其核苷酸序列与seqidno:2、3、5或6一致性至少为85%;(b)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4或7的序列一致性至少为90%;或(c)(a)或(b)核酸序列的全长互补序列;至少一个调控序列是植物中有功能的启动子。
另一方面,本发明包括含有重组dna构建体的植物或种子,其中所述重组dna构建体包含一个多核苷酸序列和与之可操作连接的至少一个调控序列,所述多核苷酸包含(a)一种多核苷酸,其核苷酸序列与seqidno:2、3、5或6的序列一致性至少为85%;(b)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4或7的序列一致性至少为90%;或者(c)(a)或(b)核苷酸序列的全长互补序列。
另一方面,本发明包含在其基因组中含有重组dna构建体的植物,所述重组dna构建体包含一个多核苷酸和与之相连的可操作连接的至少一个调控元件,其中,所述多核苷酸包含(a)一种多核苷酸,其核苷酸序列与seqidno:2、3、5或6的序列一致性至少为85%;(b)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4或7的序列一致性至少为90%;或者(c)(a)或(b)核苷酸序列的全长互补序列,其中与对照植物相比,所述转基因植物显示改变的开花时间,在所述植物中过量表达所述多核苷酸促进营养生长向生殖生长的转变,降低所述多核苷酸的表达延长营养生长向生殖生长的转变时间。
另一方面,本发明包括本发明公开的任意植物,其中植物选自水稻、玉米、大豆、向日葵、高粱、油菜、小麦、苜蓿、棉花、大麦、栗、甘蔗和柳枝稷。
另一方面,本发明还公开了调控植物开花时间的方法,其包含:(a)将重组dna构建体转入可再生植物细胞,所述重组dna构建体包含与至少一个调控序列可操作连接的多核苷酸,其中所述多核苷酸编码的多肽的氨基酸序列与seqidno:4或7相比序列一致性至少为80%;(b)由步骤(a)后的可再生细胞再生转基因植物,其中转基因植物在其基因组中包含重组dna构建体;和(c)由步骤(b)的转基因植物获得子代植物,其中所述子代植物在其基因组中包含重组dna构建体;与不含有重组dna构建体的对照植物相比,所述子代植物显示改变的开花时间。在所述植物中,过量表达所述多核苷酸促进植物营养生长向生殖生长的转变;降低所述多核苷酸的表达延长营养生长向生殖生长转变的时间。
另一方面,本发明涉及一个重组dna构建体,其包含本发明中任意分离的多聚核苷酸,并与至少一个调控序列可操作连接;以及包含重组dna构建体的细胞、植物和种子。所述细胞包括真核细胞,如酵母、昆虫或植物细胞;或原核细胞,如细菌。
附图简述和序列表
根据以下的发明详述和附图以及序列表,可更全面地理解本发明,以下的发明详述和附图以及序列表形成本申请的一部分。
图1为实时pcr分析测定的不同转基因株系叶片中osmbd1基因的相对表达水平。zh11-tc叶片中基因的表达水平设置为1.00,每个转基因株系表达量柱上面的数字表示与zh11-tc相比的变化倍数。zh11-tc为组织培养获得的zh11,zh11-wt是野生型中花11,dp0158代表转空载体dp0158的zh11。
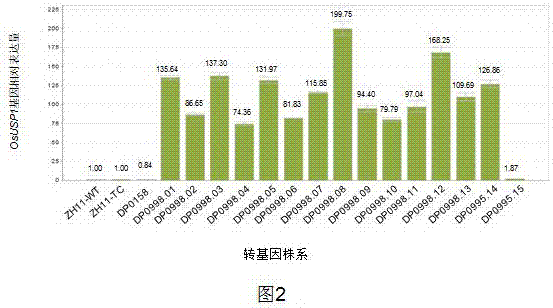
图2为实时pcr分析测定的不同转基因株系叶片中osusp1基因的相对表达水平。zh11-tc叶片中基因的表达水平设置为1.00,每个转基因株系表达量柱上面的数字表示与zh11-tc相比的变化倍数。zh11-tc为组织培养获得的zh11,zh11-wt是野生型中花11,dp0158代表转空载体dp0158的zh11。
图3为干旱试验中宁夏田间土壤体积含水量在osmbd1转基因水稻不同发育时期的变化。osmbd1转基因水稻在断水后20天开始抽穗,zh11-tc水稻在断水后43天开始抽穗。
图4为干旱试验中宁夏田间土壤体积含水量在osusp1转基因水稻不同发育时期的变化。osusp1转基因水稻在断水后22天开始抽穗,zh11-tc水稻在断水后43天开始抽穗。
表1.序列表中核苷酸和氨基酸序列编号
表2.克隆水稻花期调控基因的引物
表3.pcr反应混合液
表4.克隆水稻花期调控基因的pcr循环状况
表5.t1代osmbd1转基因水稻的开花性状、植株高度和产量(第一次北京验证)
表6.t1代osmbd1转基因水稻在载体水平上产量分析(第一次北京验证)
表7.t1代osmbd1转基因水稻在载体水平上植株高度分析(第一次北京验证)
表8.t1代osmbd1转基因水稻在载体水平上有效穗数分析(第一次北京验证)
表9.t1代osmbd1转基因水稻在海南的开花性状、植株高度和产量
表10.t1代osmbd1转基因水稻在北京的开花性状、植株高度和产量(第二次北京验证)
表11.t1代osmbd1转基因水稻在宁夏的开花性状、植株高度和产量
表12.t2代osmbd1转基因水稻在海南的开花性状、植株高度和产量
表13.t2代osmbd1转基因水稻在北京的开花性状、植株高度和产量(第一次试验)
表14.t2代osmbd1转基因水稻在北京的开花性状、植株高度和产量(第二次试验)
表15.t2代osmbd1转基因水稻在宁夏的开花性状、植株高度和产量
表16.osmbd1转基因水稻在田间干旱条件下的产量分析
表17.t1代osusp1转基因水稻的开花性状、植株高度和产量(第一次北京验证)
表18.t1代osusp11转基因水稻在载体水平上产量分析(第一次北京验证)
表19.t1代osusp1转基因水稻在载体水平上植株高度分析(第一次北京验证)
表20.t1代osusp1转基因水稻在载体水平上有效穗数分析(第一次北京验证)
表21.t1代osusp1转基因水稻在海南的开花性状、植株高度和产量
表22.t1代osusp1转基因水稻在北京的开花性状、植株高度和产量(第二次北京验证)
表23.t1代osusp1转基因水稻在宁夏的开花性状、植株高度和产量
表24.t2代osusp1转基因水稻在海南的开花性状、植株高度和产量
表25.t2代osusp1转基因水稻在北京的开花性状、植株高度和产量(第一次试验)
表26.t2代osusp1转基因水稻在北京的开花性状、植株高度和产量(第二次试验)
表27.t2代osusp1转基因水稻在宁夏的开花性状、植株高度和产量
表28.osusp1转基因水稻在田间干旱条件下的产量分析
表1.序列表中核苷酸和氨基酸序列编号
序列描述以及关联的序列表遵循如37c.f.r.§1.821-1.825中所列出的管理专利申请中的核苷酸和/或氨基酸序列公开的规则。序列表包含核苷酸序列字符的单字母码以及氨基酸的三字母码,如遵照iupac-iubmb标准所定义的,该标准在nucleicacidsres.13:3021-3030(1985)以及在biochemicalj.219(no.2):345-373(1984)中有所描述,这两篇文献以引用的方式并入本文。用于核苷酸和氨基酸序列数据的符号和格式遵循37c.f.r.§1.822中列出的规则。
seqidno:1为dp0158载体的核苷酸序列。
seqidno:2为osmbd1基因cdna的核苷酸序列。
seqidno:3为osmbd1基因cds的核苷酸序列。
seqidno:4为osmbd1蛋白的氨基酸序列。
seqidno:5为osusp1基因cdna的核苷酸序列。
seqidno:6为osusp1基因cds的核苷酸序列。
seqidno:7为osusp1蛋白的氨基酸序列。
seqidno:8为克隆osmbd1基因cdna的正向引物。
seqidno:9为克隆osmbd1基因cdna的反向引物。
seqidno:10为克隆osusp1基因cdna的正向引物。
seqidno:11为克隆osusp1基因cdna的反向引物。
seqidno:12为osmbd1基因实时rt-pcr分析的正向引物。
seqidno:13为osmbd1基因实时rt-pcr分析的反向引物。
seqidno:14为osusp1基因实时rt-pcr分析的正向引物。
seqidno:15为osusp1基因实时rt-pcr分析的反向引物。
详细说明
本文中所列出的每篇参考文献的公开内容的全文均以引用的方式并入本文。
如本文所用的并在所附权利要求书中的单数形式“一个”和“所述”包括复数涵义,除非上下文中清楚地另有指明。因此,例如,“一株植物”的涵义包括多株该类植物。“一个细胞”的涵义包括一个或多个细胞及其本领域的技术人员已知的等同物,等等。
如本文所述:
“osmbd1”是甲基结合结构域蛋白(methyl-bindingdomainprotein,mbd),涉及水稻基因位点loc_os05g33550.1编码的能够调控水稻开花性状的多肽。“mbd1多肽”此处涉及osmbd1多肽和源于其他植物的同源物。
osmbd1多肽(seqidno:4)是水稻基因位点loc_os05g33550.1的编码序列(cds)(seqidno:3)或核酸序列(seqidno:2)编码的氨基酸序列。tigr(theinternetatplantbiologymsu.edu/index.shtml)中,该多肽的注释为“甲基结合结构域蛋白mbd,推测,表达”,但是没有在先的功能介绍。
“osusp1(普遍胁迫蛋白1)”指水稻基因位点loc_os07g36600.1编码的调控水稻开花特征的水稻多肽。“usp1多肽”此处指osusp1多肽和源于其他植物的同源物。
osusp1多肽(seqidno:7)是水稻基因位点loc_os07g36600.1的编码序列(cds)(seqidno:6)或核酸序列(seqidno:5)编码的氨基酸序列)。tigr中,该多肽的注释为“含有普遍胁迫蛋白的蛋白,推测,表达”,然而没有在先的功能介绍。
本发明中的单子叶植物包括禾本科的植物;双子叶植物包括十字花科、豆科和茄科的植物。
“开花”指开花的过程,即在适宜的温度和湿度下,颖壳开裂,花药散射,或花的形成过程。此处开花用于表示从幼穗分化、成熟到幼穗抽出的过程。
“花发育”的意思是指从花分生组织开始到发育成熟花的花或花序的发育。
“生殖发育”的意思是指从花分生组织开始经过授粉到果实成熟的花或花序发育过程。
此处“早开花”的植物指开花早于对照的植物,因此这个词指的是显示开花起始较早的植物。植物的开花时间指播种到第一个花序出现的天数,例如,植物的“开花时间”可以使用wo2007/093444中描述的方法确定。
“抽穗”此处指禾谷类作物发育完全的幼穗从剑叶鞘内伸出的时期或状态。
“抽穗期”和“抽穗时间”本文可互换使用,指从种子播种50%幼穗从水稻植株剑叶鞘抽出的天数。抽穗期是重要的农业性状,由基本的营养基因和光周期敏感基因调控,在水稻品种的适应性和地理分布起关键的作用。适当的抽穗期是获得理想产量的前提。
通常情况下,水稻穗抽出后即开花,因此本文抽穗期也用于表示开花期。
“植株高度”本文指从地面到单株植株最高穗或叶顶部的高度。
“全长互补序列”是指给定核苷酸序列的互补序列,互补序列和核苷酸序列含有相同的核苷酸数,并且100%的互补。
“表达序列标签”(“est”)是得自cdna文库的dna序列,并且因此是已经被转录的序列。est通常通过cdna插入序列单程测序获取。将完整的cdna插入序列称为“全长插入序列”(“fis”)。“重叠群”序列是由选自但不限于est、fis和pcr序列的两个或更多个序列装配成的序列。将编码完整或功能性蛋白的序列称为“完全基因序列”(“cgs”),该序列可得自fis或重叠群。
“转基因的”指其基因组因异源核酸(如重组dna构建体)的存在而发生改变的任何细胞、细胞系、愈伤组织、组织、植物部分或植物,包括那些最初的转基因事件以及从最初的转基因事件通过有性杂交或无性生殖而产生的那些。如本文所用的术语“转基因”不涵盖通过常规植物育种方法或通过诸如随机异花受精、非重组病毒感染、非重组细菌转化、非重组转座或自发突变之类的自然发生事件导致的基因组(染色体基因组或染色体外基因组)改变。
“对照”、“对照植物”或“对照植物细胞”为测定受测试植物或植物细胞的表型变化提供参考,由于转化,受测植物或植物细胞的基因组改变影响到目的基因,测试的植物或植物细胞可以是转基因植物或转基因植物细胞的子代。
对照植物或对照植物细胞包括,例如:(a)用于基因改变产生受测植物或细胞的野生型植物或细胞;(b)相同基因组作为起始材料但是转入空载体(如带有标记基因的并对目标的性状没有影响的载体)的植物或植物细胞;(c)转基因植物或植物细胞性状分离,获得的非转基因的子代植物或植物细胞;(d)没有暴露于可以诱导基因表达的条件或刺激下的,与转基因植物或植物细胞基因组相同的植物或植物细胞;(e)特定的目标基因不表达情况下的,转基因植物或者植物细胞本身。
本文中,zh11-wt、zh11-tc和空载体植物指对照植物。zh11-wt代表野生型中花11,zh11-tc代表通过组织培养中花11获得的水稻植株,空载体代表转化空载dp0158获得水稻植株。
“基因组”在用于植物细胞时不仅涵盖存在于细胞核中的染色体dna,而且还包括存在于细胞的亚细胞组分(如线粒体、质粒)中的细胞器dna。
“植物”包括整个植株、植物器官、植物组织、种子和植物细胞以及同一植株的子代。植物细胞包括但不限于得自下列物质的细胞:种子、悬浮培养物、胚、分生区域、愈伤组织、叶、根、芽、配子体、孢子体、花粉和小孢子。
“子代”包括植物的任何后续世代。
“转基因植物”包括在其基因组内包含异源多核苷酸的植物。例如异源多核苷酸能够稳定地整合进基因组中,并遗传连续的世代。异源多核苷酸可单独地或作为重组dna构建体的部分整合进基因组中。t0植物直接源于转化和再生过程,t0植物的子代为t1代(第一个子代),t2代(第二个子代)等。
针对序列而言的“异源”意指来自外来物种的序列,或者如果来自相同物种,则指通过蓄意的人为干预而从其天然形式发生了组成和/或基因座的显著改变的序列。
“多核苷酸”、“核酸序列”、“核苷酸序列”或“核酸片段”可互换使用并且是任选含有合成的、非天然的或改变的核苷酸碱基的单链或双链rna或dna聚合物。核苷酸(通常以它们的5′-单磷酸形式存在)通过如下它们的单个字母名称来指代:“a”为腺苷酸或脱氧腺苷酸(分别对应rna或dna),“c”表示胞苷酸或脱氧胞苷酸,“g”表示鸟苷酸或脱氧鸟苷酸,“u”表示尿苷酸,“t”表示脱氧胸苷酸,“r”表示嘌呤(a或g),“y”表示嘧啶(c或t),“k”表示g或t,“h”表示a或c或t,“i”表示肌苷,并且“n”表示任何核苷酸。
“多肽”、“肽”、“氨基酸序列”和“蛋白质”在本文中可互换使用,指氨基酸残基的聚合物。该术语适用于其中一个或多个氨基酸残基是相应的天然存在的氨基酸的人工化学类似物的氨基酸聚合物,以及适用于天然存在的氨基酸聚合物。术语“多肽”、“肽”、“氨基酸序列”和“蛋白质”还可包括修饰形式,包括但不限于糖基化、脂质连接、硫酸盐化、谷氨酸残基的γ羧化、羟化和adp-核糖基化。
“信使rna(mrna)”指无内含子并且可通过细胞翻译成蛋白质的rna。
“cdna”指与mrna模板互补并且利用逆转录酶从mrna模板合成的dna。cdna可为单链的或者可用dna聚合成酶i的klenow片段转化成双链形式。
“成熟”蛋白质指经翻译后加工的多肽;即已经去除了存在于初级翻译产物中的任何前肽或原肽的多肽。
“前体”蛋白质指mrna的翻译初级产物;即带有前肽和原肽的蛋白质。前肽和原肽可为并且不限于细胞内定位信号。
“分离的”指物质,例如核酸和/或蛋白质,该物质基本上不含在天然存在的环境中通常伴随该物质或与其反应的组分,或者说是该物质被从所述组分移出。分离的多核苷酸可从它们天然存在的宿主细胞中纯化。技术人员已知的常规核酸纯化方法可用于获得分离的多核苷酸。该术语也涵盖重组多核苷酸和化学合成的多核苷酸。
“重组体”指(例如)通过化学合成或者通过用基因工程技术操纵分离的核酸片段来实现的两个原本分离的序列片段的人工组合。“重组体”也包括指已经通过引入异源核酸而进行了修饰的细胞或载体,或源于经这样修饰的细胞的细胞,但不涵盖由天然发生的事件(如自发突变、自然转化/转导/转座)对细胞或载体的改变,例如没有蓄意人为干扰而发生的那些。
“非基因组核酸序列”、“非基因组核酸分子”或“非基因组多核苷酸”指与天然或基因组核酸序列相比,存在一处或多处核酸序列改变的核酸分子。在某些实施方案中,天然或基因组核酸分子的改变包括但不限于:由于遗传密码的简并性而产生的核酸序列变化;植物表达的核酸序列密码子优化;与天然或基因组序列相比,至少一个氨基酸的替代、插入、删除和/或添加而引起的核酸序列的变化;与基因组核酸序列相连的一个或多个内含子的移除;一个或多个异源内含子的插入;与基因组核酸序列相连一个或多个上游或下游调控区域的删除,一个或多个异源上游或下游调控区域的插入;与基因组核酸序列相连的5’和/或3’非翻译区的删除;一个异源5’和/或3’非翻译区的插入;和多聚腺苷酸位点的修饰。在一些实施方案中,非基因组核酸分子是cdna。在一些实施方案中,非基因组核酸分子为合成的核酸序列。
“重组dna构建体”指在自然界中通常不会一起存在的核酸片段的组合。因此,重组dna构建体可包含源于不同来源的调控序列和编码序列,或源于相同来源但以不同于通常天然存在的方式排列的调控序列和编码序列。
术语“入门克隆”和“入门载体”本文可互换使用。
“调控序列”和“调控元件”可互换使用,指位于编码序列的上游(5′非编码序列)、中间或下游(3′非编码序列),并且影响相关编码序列的转录、rna加工或稳定性或者翻译的核苷酸序列。调控序列可包括但不限于启动子、翻译前导序列、内含子和多腺苷酸化识别序列。
“启动子”指能够控制另一核酸片段转录的核酸片段。
“植物启动子功能”是能够控制植物细胞中的转录的启动子,无论其是否来源于植物细胞。
“组织特异性启动子”和“组织优选启动子”可互换使用,并且指主要但非必须专一地在一种组织或器官中表达,而是也可在一种特定细胞中表达的启动子。
“发育调控启动子”指其活性由发育事件决定的启动子。
术语“可操作地连接”指核酸片段连接成单一片段,使得其中一个核酸片段的功能受到另一个核酸片段的调控。例如,在启动子能够调节核酸片段的转录时,该启动子与该核酸片段进行了可操作地连接。
“表达”指功能产物的产生。例如,核酸片段的表达可指核酸片段的转录(如转录生成mrna或功能rna)和/或rna翻译成前体或成熟蛋白质。
“表型”意指细胞或生物体的可检测的特征。
有关将核酸片段(例如重组dna构建体)插入细胞内的“导入”是指“转染”或“转化”或“转导”,并且包括指将核酸片段整合进真核或原核细胞中,在该细胞中核酸片段可整合进细胞的基因组(如染色体、质粒、质体或线粒体dna)内,转变成自主的复制子或瞬时表达(如转染的mrna)。
“转化细胞”是将核酸片段(如重组dna构建体)导入其中的任何细胞。
本文所用的“转化”指稳定转化和瞬时转化两者。
“稳定转化”指将核酸片段导入宿主生物体的基因组中,导致基因稳定遗传。一旦稳定转化,核酸片段稳定地整合进宿主生物体和任何连续世代的基因组中。
“瞬时转化”指将核酸片段导入宿主生物体的核中或包含dna的细胞器中,引起基因表达而没有基因稳定遗传。
“等位基因”是占据染色体上给定位点的基因的几种供选择形式的其中一种。当二倍体植物中一对同源染色体上给定基因座上存在的等位基因相同时,该植物在该基因座处是纯合的。如果二倍体植物中一对同源染色体上给定基因座上存在的等位基因不同,则该植物在该基因座处是杂合的。如果转基因存在于二倍体植物中一对同源染色体中的其中之一上,则该植物在该基因座处是半合子的。
“叶绿体转运肽”是与蛋白协同翻译并将蛋白导向叶绿体或翻译蛋白的细胞中存在的其它质体类型的氨基酸序列。“叶绿体转运序列”指编码叶绿体转运肽的核苷酸序列。“信号肽”是一种与蛋白协同翻译并将蛋白导向分泌器官的氨基酸序列(chrispeels,m.(1991)ann.rev.plantphys.plantmol.biol.42:21-53)。如果将所述蛋白导向液泡,可另外加上液泡靶向信号,或者如果将所述蛋白导向内质网,可加上内质网驻留信号。如果将蛋白导向细胞核,将移除任何存在的信号肽并用以核定位信号替代(raikhel(1992)plantphys.100:1627-1632)。“线粒体信号肽”是引导前体蛋白进入线粒体的氨基酸序列(zhang和glaser(2002)trendsplantsci7:14-21)。
dna核酸酶和其他的突变酶结构域可以与dna结合结构域融合在靶dna上产生双链断裂(dsb),dna结合结构域包括,例如一个特异dna结合结构域,或一个位点特异dna结合结构域。位点特异dna结合结构域包括但不限于tal(转录激活样效应因子)或锌指结合结构域。
dna结合结构域与dna核酸酶融合的例子包括但不限于talen和多talen。转录激活样效应因子核酸酶(talens)是tal效应因子dna结合结构域融合到dna酶的结构域产生的人工限制性酶。tal蛋白是细菌产生的,包括高度保守的33~34氨基酸dna结合结构域序列(pct公开号wo2014127287,美国专利公开号us20140087426)。
起初的talen嵌合体是由野生型的foki核酸内切酶结构域制备的,而talen也包括源于foki核酸内切酶结构域变异提高了切割特异性和切割活性突变嵌合体。在一些例子中,多个talen可以靶向多个基因区域表达。
锌指是另一种类型的dna结合结构域,可以将突变到引入靶dna。
各种蛋白工程技术可以用于改变锌指dna结合特异性,这样设计的锌指的串联重复序列可用于靶向期望的基因组dna序列。融合第二个蛋白结构域例如转录抑制子到锌指,可以与附近的yep基因启动子结合,从而降低mbd1和usp1基因表达水平。
在一个实施例中,一个调控元件驱动内源基因表达或编码序列本身,例如,可能会通过crispr/cas9体系的基因组编辑技术被编辑或插入植物中。
crispr位点(成簇的规律间隔短回文重复,也被称作spidrs,间隔穿插的顺向重复)构成近来描述的dna位点家族,crispr位点由短且高度保守的dna重复(典型的24~40bp,重复1~140次,也称为crispr重复)组成,crispr位点部分回文。重复序列(通常具有品种特异性)被固定长度(代表性的20~58bp,根据crispr位点而不同)的可变序列间隔开(公布与2007年3月1日的wo2007/025097)。
cas基因指通常耦合的,关联或邻近crispr位点或在crispr位点侧翼的附近的基因。术语“cas基因”、“crispr关联(cas)基因”此处可以互换使用。(haft等(2005),计算生物学,ploscomputbiol1(6):e60,doi:10.1371/journal.pcbi.0010060)。如本文所描述,41个crispr关联(cas)基因家族进行了描述,除了四个已知基因家族。这表明crispr系统属于不同的类别,具有不同的重复模式,基因组别和物种范围。一个给定的crispr位点不同物种间的cas基因数目不同。
cas核酸内切酶指cas基因编码的cas蛋白,其中,所述cas蛋白可以向靶dna序列导入双链断裂。cas核酸切酶由引导多聚核苷酸引导识别和选择性的向细胞基因组特定靶位点导入双链断裂(u.s.2015/0082478)。引导多聚核苷酸/cas核酸内切酶体系包括cas核酸内切酶和引导多聚核苷酸组成的复合体,引导多聚核苷酸可以向dna靶序列导入双链断裂。如果一个正确的前间区序列邻近基序(pam)导向的靶序列的3’端,cas核酸内切酶解开基因组靶位点邻近dna双链,引导rna切割靶序列dna双链的识别位点。cas核酸内切酶可以通过本领域已知的方法直接导入细胞,例如,但不限于瞬时导入方法,转染和/或局部适用。
如本文中使用,术语“引导rna”指两个rna分子合成的融合,crrna(crisprrna)包括一个可变的靶结构域,和一个tracrrna。在一个实施例中,引导rna包括一个12~30bp核苷酸序列的可变的靶结构域和可以与cas核酸内切酶相互作用的rna片段。
如本文所述,术语“引导多聚核苷酸”指可以与cas核酸内切酶构成复合体的多聚核苷酸序列,可以使cas核酸内切酶识别并随意切割dna靶位点(u.s.2015/0082478)。引导多聚核苷酸可以是单链分子也可以是双链分子,可以是rna序列,dna序列或它们的组合(rna-dna组合序列)。可选的,引导多聚核苷酸可以包括至少一个核苷酸,磷酸二酯键或修饰键,例如,但不限于锁定核苷酸(lna)、5-甲基脱氧胞苷、2,6-二氨基嘌呤、2-氟腺苷,2-氟脲苷、2-甲基rna、硫代磷酸酯键,一种胆固醇分子链、聚乙二醇分子链、以间隔18(六乙二醇链)分子链、或5至3共价键产生的环。引导多聚核苷酸仅包括脱氧核糖核酸的也被称作”引导rna”。
术语“可变靶结构域”或“vt结构域”此处可互换使用,包括与双链dna靶位点的一条单链(核苷酸序列)互补的核苷酸序列。第一个核苷酸序列结构域(vt结构域)与靶序列的互补百分数至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、63%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%。可变靶结构域的长度至少为12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29或者30bp的核苷酸。在一些实施例中,可变靶结构域由邻近延伸的12~30bp核苷酸组成。可变靶结构域是由dna序列、rna序列、修饰的dna序列、修饰的rna序列或它们的任意组合组成。
术语“cas核酸内切酶识别结构域”或引导多聚核苷酸的“cer结构域”此处可互换使用,包括可以与cas核酸内切酶多肽相互作用的核苷酸序列(诸如引导多聚核苷酸的第二核苷酸序列)。cer结构域由dna序列、rna序列、修饰的dna序列、修饰的rna序列(例如此处描述的修饰)或它们的任意组合构成。
核苷酸序列连接单引导多聚核苷酸的crnucleotide和tracrnucleotide可以组成rna序列、dna序列、,或rna-dna组合序列。在一个实施例中,核苷酸序列连接单引导多聚核苷酸的crnucleotide和tracrnucleotide组成长度至少为3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100bp的核苷酸序列。在另一个实施例中,核苷酸序列连接单多聚核苷酸的crnucleotide和tracrnucleotide组成四核苷酸环序列,诸如,但不限于gaaa四核苷酸环序列。
序列比对和同一性百分比可用设计用于检测同源序列的多种比较方法来测定,这些方法包括但不限于生物信息计算包(inc.,madison,wi)的程序。除非另外说明,本文提供的序列的多重比对用clustalv比对方法(higgins和sharp,1989,cabios.5:151-153)采用默认参数(空位罚分=10,空位长度罚分=10)执行。用clustalv方法进行成对比对和蛋白质序列的同一性百分比计算的默认参数为ktuple=1、空位罚分(gappenalty)=3、窗口(window)=5并且diagonalssaved=5。而对于核酸,这些参数为ktuple=2,空位罚分=5,窗口=4并且diagonalssaved=4。用clustalv程序比对序列后,可通过查看同一程序中的“序列距离”表来获得“同一性百分比”和“趋异”值。除非另外说明,本文提供的和申明的同一性百分比和趋异度是以该方式计算。
本文使用的标准重组dna和分子克隆技术是本领域所熟知的并且在如下文献中有更全面的描述:sambrook,j.,fritsch,e.f.和maniatis,t.,molecularcloning:alaboratorymanual;coldspringharborlaboratorypress:coldspringharbor,1989(下文称为“sambrook”)。
现在转向实施方案:
实施方案包括分离的多核苷酸和多肽、用于调控植物开花时间的重组dna构建体、包含这些重组dna构建体的组合物(例如植物或种子),以及利用这些重组dna构建体的方法。
分离的多核苷酸和多肽:
本发明包括如下分离的多核苷酸和多肽:
在一些实施方案中,多核苷酸编码mbd1或usp1多肽。
在一些实施方案中,分离的多核苷酸包含(i)编码多肽的核酸序列,所述多肽具有的氨基酸序列在与seqidno:4或7进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(ii)(i)的核酸序列的全长互补序列,其中全长互补序列和(i)的核酸序列由相同数目的核苷酸组成并且是100%互补的。任一上述分离的多核苷酸可用于本发明的任何重组dna构建体。
在一些实施方案中,分离的多肽,其氨基酸序列在与seqidno:4或7进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性。所述多肽优选地为花期调控多肽mbd1或usp1。
在一些实施方案中,分离的多核苷酸,其包括:(i)核酸序列,所述核酸序列在与seqidno:2、3、5或6进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(ii)(i)的核酸序列的全长互补序列。任一上述分离的多核苷酸可用于本发明的任何重组dna构建体。分离的多核苷酸优选为编码花期调控蛋白。过量表达该花期调控多肽促进植物从营养生长到生殖生长转变;减少该多肽的表达量延长了从营养生长到生殖生长的转变时间。
重组dna构建体和抑制dna构建体
在一方面,本发明包括重组dna构建体和抑制dna构建体。
在一个实施方案中,重组dna构建体包含可操作地连接至少一种调控序列(如,在植物中有功能的启动子)的多核苷酸,其中所述多核苷酸包括(i)核酸序列,所述核酸序列编码的氨基酸序列与seqidno:4或者7进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%的序列一致性;或(ii)(i)的核酸序列的全长互补序列。
在另一个实施方案中,重组dna构建体包含可操作地连接至少一种调控序列(如,在植物中有功能的启动子)的多核苷酸,其中所述多核苷酸包括(i)核酸序列,所述核酸序列在与seqidno:2、3、5或6进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%的序列一致性;或(ii)(i)的核酸序列的全长互补序列。
在另一实施方案中,重组dna构建体包含可操作地连接至少一种调控序列(如,在植物中有功能的启动子)的多核苷酸,其中所述多核苷酸编码mbd1或usp1。这些多肽具有调控花期的活性,并可来自,例如水稻(oryzasativa)、野生稻(oryzaaustraliensis)、短舌野生稻(oryzabarthii)、非洲型稻(oryzaglaberrima)、阔叶稻(oryzalatifolia)、长雄野生稻(oryzalongistaminata)、南方野生稻(oryzameridionalis)、药用野生稻(oryzaofficinalis)、斑点野生稻(oryzapunctata)、普通野生稻(oryzarufipogon)(红稻)、印度野生稻(oryzanivara)、拟南芥(arabidopsisthaliana)、玉米(zeamays)、大豆(glycinemax)、烟豆(glycinetabacina)、野大豆(glycinesoja)和短绒野大豆(glycinetomentella)。
在另一方面,本发明包括抑制dna构建体。
抑制dna构建体包括至少一个调控序列(如植物中有功能的启动子)可操作的连接至全部或部分如下序列:(i)核酸序列,所述核酸序列编码的多肽,其氨基酸序列与seqidno:4或7的序列一致性至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%,或者(ii)(a)(i)核酸序列的全长互补序列;或者(b)源于目标基因的正义链或反义链的全部或部分的区域,所述区域的核酸序列与该目标基因的正义链或反义链的全部或部分相比具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%得序列一致性,其中所述目标基因编码花期调控多肽mbd1或usp1;或者(c)全部或部分(i)与seqidno:2或5的核酸序列一致性至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%,或者(ii)与seqidno:3或6的核酸序列一致性至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%,或者(iii)(c)(i)或者(c)(ii)的全长互补核酸序列。抑制dna构建体包括共抑制构建体、反义构建体、病毒抑制构建、发夹抑制构建体、茎-环抑制构建体、双链rna产生构建体、rnai构建体或小rna构建体(例如sirna构建体或mirna构建体)。
应当理解(正如本领域技术人员将会理解的),本发明不仅仅涵盖这些具体的示例性序列。导致给定位点处产生化学上等价的氨基酸但不影响所编码多肽的功能特性的核酸片段中的改变是本领域众所周知的。例如,丙氨酸(一种疏水性氨基酸)的密码子可被编码另一个疏水性较弱的残基(例如甘氨酸)或疏水性较强的残基(例如缬氨酸、亮氨酸或异亮氨酸)的密码子取代。类似地,导致一个带负电荷的残基替换为另一个带负电荷的残基(例如,天冬氨酸替代谷氨酸)或者一个带正电荷的残基替换为另一个带正电荷的残基(例如,赖氨酸替换精氨酸)的改变也可预期产生功能上等价的产物。导致多肽分子的n-末端和c-末端部分改变的核苷酸变化也将预计不会改变多肽的活性。所提出的修饰中的每一种均完全在本领域常规技术内,如测定所编码的产物的生物活性的保留。
“抑制dna构建体”是在转化或稳定整合进植物基因组时,导致该植物中的靶基因“沉默”的重组dna构建体。对该植物来说,该靶基因可为内源性的或是转基因的。如本文针对靶基因所使用的,“沉默”通常指在由靶基因表达的mrna或蛋白质/酶的水平上的抑制,和/或在酶活性或蛋白质功能性的水平上的抑制。本文中可交换使用的术语“抑制”、“抑制性”以及“沉默”包括降低、减少、减退、减小、抑制、消除或防止。“沉默”或“基因沉默”不特指机理并且包括但不限于反义、共抑制、病毒-抑制、发夹抑制、茎-环抑制、基于rnai的方法以及基于小rnai的方法。
抑制dna构建体可包含源自所关注的靶基因的区域并且可包含所关注的靶基因的有义链(或反义链)的核酸序列的全部或部分。取决于所要利用的方法,该区域可与所关注基因的有义链(或反义链)的全部或部分100%相同或者具有少于100%序列的一致性(如,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的一致性)。
抑制dna构建体是本领域所熟知的,一旦选定所关注的靶基因就很容易构建,并且包括但不限于共抑制构建体、反义构建体、病毒-抑制构建体、发夹抑制构建体、茎-环抑制构建体、产生双链rna的构建体,以及更通常的是,rnai(rna干扰)构建体和小rna构建体,例如sirna(短干扰rna)构建体和mirna(微rna)构建体。
“反义抑制”指产生能够抑制靶基因或基因产物表达的反义rna转录物。“反义rna”指与靶初级转录物或mrna的全部或部分互补,并阻断分离的靶核酸片段表达的rna转录物(美国专利号5,107,065)。反义rna可与特定基因转录物的任何部分,即5′非编码序列、3′非编码序列、内含子或编码序列互补。
“共抑制”指产生能够抑制靶基因或基因产物表达的有义rna转录物。“有义”rna指包括mrna并且可在细胞内或体外翻译成蛋白质的rna转录物。此前,已通过着眼于以有义方向过表达与内源mrna具有同源性的核酸序列(其导致与过表达的序列具有同源性的所有rna减少)设计出了植物中的共抑制构建体(参见vaucheret等人,plantj.,16:651-659(1998);以及gura,nature404:804-808(2000))。
另一种变型描述了将植物病毒序列用于引导对近端mrna编码序列的抑制(于1998年8月20日公开的pct公开wo98/36083)。
rna干扰(rnai)是指由短干扰性rna(sirna)介导的动物中序列特异性转录后基因沉默的过程(fire等人,nature391:8061998)。在植物中的对应过程通常称为转录后基因沉默(ptgs)或rna沉默,并且在真菌中也称为阻抑作用(quelling)。据信转录后基因沉默过程是用于防止外来基因表达的进化保守性细胞防御机制,并且通常由不同植物区系和门所共有(fire等人,trendsgenet.15:358(1999))。
小rna在控制基因表达中起重要作用。很多发育过程(包括开花)的调节是由小rna控制的。现在有可能通过使用在植物中产生小rna的转基因构建体来以工程手段改变植物基因的基因表达。
小rna似乎是通过与互补rna或dna靶序列碱基配对来行使功能的。当与rna结合时,小rna或者引发靶序列的rna裂解或者引发翻译抑制。当与dna靶序列结合时,据信小rna可介导靶序列的dna甲基化。无论具体机制是什么,这些事件的后果是基因表达受到抑制。
微rna(mirna)是长度为约19至约24个核苷酸(nt)的已经在动物和植物中鉴定出的非编码rna(lagos-quintana等人,science294:853-8582001,lagos-quintana等人,curr.biol.12:735-739(2002);lau等人,science294:858-862(2001);lee和ambros,science294:862-864(2001);llave等人,plantcell14:1605-1619(2002);mourelatos等人,genes.dev.16:720-728(2002);park等人,curr.biol.12:1484-1495(2002);reinhart等人,genes.dev.16:1616-1626(2002))。它们是由大小为大约70至200nt较长的前体转录物加工生成的,并且这些前体转录物能够形成稳定的发夹结构。
微rna(mirna)看起来通过与位于由这些基因产生的转录物中的互补序列结合来调节靶基因。看起来有可能mirna可进入至少两条靶基因调控途径:(1)翻译抑制;和(2)rna裂解。进入rna裂解途径的微rna类似于在动物中的rna干涉作用(rnai)和植物中的转录后基因沉默(ptgs)期间生成的21-25nt的短干涉rna(sirna),并且可能掺入了rna诱导沉默复合物(risc),该复合物与rnai相似或相同。
调控序列:
本发明的重组dna构建体(包括抑制dna构建体)可包含至少一种调控序列。
调控序列可为启动子或增强子。
多种启动子可用于本发明的重组dna构建体中。可根据所需结果来选择启动子,并且可包括用于在宿主生物体中表达的组成型启动子、组织特异性启动子、诱导型启动子或其他启动子。
一般将引起基因在多数细胞类型中在多数情况下表达的启动子称为“组成型启动子”。
虽然候选基因当通过组成型启动子驱动表达时可预测其效应,但候选基因在35s或ubi启动子控制下的高水平、组成型表达可具有多重效应。使用组织特异和/或胁迫特异启动子可消除不需要的效应但保留调控植物开花时间的能力。在拟南芥属中已经观察到了该效应(kasuga等人(1999)naturebiotechnol.17:287-91)。
适用于植物宿主细胞的组成型启动子包括(例如)rsyn7启动子的核心启动子和在wo99/43838和美国专利6,072,050中公开的其他组成型启动子;camv35s核心启动子(odell等人,nature313:810-812(1985));稻肌动蛋白(mcelroy等人,plantcell2:163-171(1990));泛素启动子(christensen)等人,plantmol.biol.12:619-632(1989)和christensen等人,plantmol.biol.18:675-689(1992));pemu(last等人,theor.appl.genet.81:581-588(1991));mas(velten等人,emboj.3:2723-2730(1984));als启动子(美国专利5,659,026)等。其他组成型启动子包括例如在美国专利5,608,149、5,608,144、5,604,121、5,569,597、5,466,785、5,399,680、5,268,463、5,608,142和6,177,611中公开的那些启动子。
在选择启动子用于本发明方法时,可能有利的是使用组织特异性启动子或发育调节启动子。
组织特异性启动子或发育调节启动子是这样的dna序列,其调节dna序列选择性地在对雄穗发育、结籽或两者重要的植物细胞/组织中表达,并限制这种dna序列只在植物的雄穗发育或种子成熟期间表达。任何引起所需时空表达的可鉴定启动子均可用于本发明的方法中。
种子或胚特异性的并且可用于本发明的启动子包括大豆kunitz胰蛋白酶抑制剂(kti3,jofuku和goldberg,plantcell1:1079-1093(1989))、patatin(马铃薯块茎)(rocha-sosa,m.等人,1989,emboj.8:23-29)、convicilin、豌豆球蛋白和豆球蛋白(豌豆子叶)rerie,w.g.等人,1991,mol.gen.genet.259:149-157;newbigin,e.j.等人,1990,planta180:461-470;higgins,t.j.v.等人,1988,plant.mol.biol.11:683-695)、玉米蛋白(玉米胚乳)(schemthaner,j.p.等人,1988,emboj.7:1249-1255)、菜豆蛋白(菜豆子叶)(segupta-gopalan,c.等人,1985,proc.natl.acad.sci.u.s.a.82:3320-3324)、植物血球凝集素(菜豆子叶)(voelker,t.等人,1987,emboj.6:3571-3577)、b-伴球蛋白和大豆球蛋白(大豆子叶)(chen,z-l等人,1988,emboj.7:297-302)、谷蛋白(稻胚乳)、大麦醇溶蛋白(大麦胚乳)(marris,c.等人,1988,plantmol.biol.10:359-366)、麦谷蛋白和麦醇溶蛋白(小麦胚乳)(colot,v.等人,1987,emboj.6:3559-3564)和甘薯贮藏蛋白(sporamin)(甘薯块根)(hattori,t.等人,1990,plantmol.biol.14:595-604)。可操作地连接至嵌合基因构建体异源编码区的种子特异性基因的启动子在转基因植物中保持它们的时空表达模式。这样的实施例包括在拟南芥和甘蓝型油菜(brassicanapus)种子中表达脑啡肽的拟南芥属2s种子储藏蛋白基因启动子(vanderkerckhove等人,bio/technology7:l929-932(1989))、表达荧光素酶的菜豆凝集素和β-菜豆蛋白启动子(riggs等人,plantsci.63:47-57(1989)),以及表达氯霉素乙酰转移酶的小麦谷蛋白启动子(colot等人,emboj6:3559-3564(1987))。
可诱导启动子响应内源性或外源性刺激的存在,例如,通过化合物(化学诱导剂),或响应环境、激素、化学信号和/或发育信号而选择性表达可操纵连接的dna序列。可诱导的或受调控的启动子包括(例如)受光、热、胁迫、水涝或干旱、植物激素、创伤或诸如乙醇、茉莉酮酸酯、水杨酸或安全剂之类的化学品调控的启动子。
用于本发明的启动子包括以下启动子:1)胁迫诱导型rd29a启动子(kasuga等人,1999,naturebiotechnol.17:287-91);2)大麦启动子b22e;b22e的表达是发育中的玉米籽粒中的柄所特异性的(“primarystructureofanovelbarleygenedifferentiallyexpressedinimmaturealeuronelayers(在未成熟糊粉层中差异表达的新大麦基因的一级结构)”。klemsdal,s.s.等人,mol.gen.genet.228(1/2):9-16(1991));和3)玉米启动子,zag2(“identificationandmolecularcharacterizationofzag1,themaizehomologofthearabidopsisfloralhomeoticgeneagamous”,schmidt,r.j.等人,plantcell5(7):729-737(1993);“structuralcharacterization,chromosomallocalizationandphylogeneticevaluationoftwopairsofagamous-likemads-boxgenesfrommaize”,theissen等人,gene156(2):155-166(1995);ncbigenbank登录号x80206))。zag2转录物可在授粉前5天至授粉后(dap)7至8天被检测到,并且引导ciml在发育中的雌花序的心皮中表达,ciml对发育中的玉米籽粒的籽仁而言是特异性的。ciml转录物在授粉前4至5天至授粉后6至8天被检测到。其他可用的启动子包括可源自其表达与发育中的雌小花母系相关的基因的任何启动子。
对于在发育种子组织中表达的多聚核苷酸,特殊的启动子包括种子优选的启动子,尤其是早期籽粒/胚胎启动子和晚期籽粒/胚乳启动子,授粉后籽粒的发育大致可以分为三个基本阶段,籽粒生长的停滞期起始于授粉后0天到10-12天,在此期间,籽粒不再明显的生长,但是决定籽粒活力的重要事件将在此期间发生(如细胞建成数目)。线性籽粒灌浆期起始于授粉后的10-12天并延续到授粉后的40天左右。在籽粒发育期间,籽粒达到最终的质量,并产生多种储藏物质如淀粉、蛋白质和油等;最终的成熟期起始于授粉后大约40天到收获,籽粒发育的这一过程中,籽粒开始休眠,变干。本发明中的“早期籽粒/胚乳启动子”指主要在种子发育的停滞期(也就是授粉第0天到授粉后第12天期间)驱动基因表达的启动子;“后期籽粒/胚乳启动子”主要驱动基因在授粉后12天到成熟过程中的种子中表达;表达窗口可能会有一些重叠,将根据使用的aba偶联的序列和期望的表型选择启动子。
早期籽粒/胚胎启动子包括,cim1,授粉后第5天活跃在特定组织中(wo00/11177);其它的早期籽粒/胚胎启动子包括种子偏爱启动子end1,在授粉后7-10天表达,和end2,授粉后9-14天在整个籽粒中表达,在授粉后10天在胚乳和果皮中表达(wo00/12733)。本发明中的特定方法使用的其它早期籽粒/胚乳启动子包括种子偏爱启动子ltp2(美国专利号5,525,716);玉米zm40启动子(美国专利号6,403,862);玉米nuc1c(美国专利号6,407,315);玉米ckx1-2启动子(美国专利号6,921,815和美国专利申请公开号2006/0037103);玉米lec1启动子(美国专利号7,122,658);玉米esr启动子(美国专利号7,276,596);玉米zap启动子(美国专利申请公开号20040025206和20070136891);玉米启动子eep1(美国专利申请公开号20070169226);和玉米启动子adf4(美国专利申请号60/963,878,2007年8月7日申请)。
本发明中调控核酸序列在植物中表达的其它启动子是茎特异启动子,包括苜蓿s2a启动子(genbank登录号ef030816;abrahams等(1995)plantmol.biol.27:513-528)和s2b启动子(genbank登录号ef030817)和类似的启动子。
启动子可以整个源于天然基因,或者由源于不同的天然存在的启动子的不同元件组成,或者甚至包括合成的dna片段。
用于本发明的启动子包括:rip2、mlip15、zmcor1、rab17、camv35s、rd29a、b22e、zag2、sam合成酶、泛素、camv19s、nos、adh、蔗糖合成酶、r-等位基因、维管组织优选启动子s2a(genbank登录号ef030816)和s2b(genbank登录号ef030817)及来自玉米的组成型启动子gos2。其他启动子包括根优选的启动子,例如玉米nas2启动子、玉米cyclo启动子(us2006/0156439,公开于2006年7月13日)、玉米rootmet2启动子(wo05063998,公开于2005年7月14日)、crlbio启动子(wo06055487,公开于2006年5月26日)、crwaq81(wo05035770,公开于2005年4月21日)和玉米zrp2.47启动子(ncbi登录号:u38790;gino.1063664)。
本发明的重组dna构建体也可包括其他调控序列,包括但不限于翻译前导序列、内含子和多腺苷酸化识别序列。在本发明的另一个实施方案中,本发明的重组dna构建体还包括增强子或沉默子。
内含子序列可加至5’非翻译区、蛋白编码区或3’非翻译区以增加积聚在胞浆中的成熟信息的量。已经显示,在植物和动物两者的表达构建体的转录单位中包含可剪接内含子可使基因表达在mrna和蛋白质水平上均增强高达1000倍。参见buchman和berg,mol.cellbiol.8:4395-4405(1988);callis等人,genesdev.1:1183-1200(1987)。
增强子或增强子元件指的是一个顺式作用的转录调控元件,即顺式元件,它可调控多核苷酸序列整体表达模式的一个方面,但通常不足以单独驱动转录可操作连接的多核苷酸序列。分离的增强子元件可以与一个启动子融合形成一个嵌合的启动子顺式元件,从而调节基因表达。本领域的技术人员均知道增强子,增强子包括sv40增强子区域、camv35s增强子元件等。一些增强子也可以改变普通调控元件的表达模式,例如,当没有增强子时,导致调控元件组成型表达,当存在增强子时,同一调控元件在某一特定组织或某些特定组织表达。camv35s启动子重复上游区域显示出大约增强10倍的表达量(kay,r.等,(1987)science236:1299-1302)。
本发明中使用的增强子包括camv35s(benfey等,(1990)emboj.9:1685-96)、4xb3p-camv35s增强子区域—四拷贝串联b3区域(208到155)(美国专利号5,097,025)、4xas-1p-camv35s增强子区域-四拷贝串联激活序列(83到62)(美国专利号5,097,025)、2xb1-b2p-camv35s增强子区域—二拷贝串联b1-b2区域(148到90)(美国专利号5,097,025)、2xa1-b3p-camv35s增强子区域—二拷贝串联a1-b3区域(208到46)(美国专利号5,097,025)、2xb1-b5p-camv35s增强子区域—二拷贝b1-b5区域(343到90)(美国专利号5,097,025)、欧米伽增强子或欧米伽基本增强子(gallie等(1989)molecularbiologyofrnaed.cech(liss,newyork)237-256和gallie等(1987)gene60:217-25)、美国专利号7,803,992的增强子和甘蔗棒状病毒(scbv)增强子元件(wo2013130813)。
任何植物都可以选择用来鉴定将用于本发明重组dna构建体的调控序列和基因。适用于分离基因和调控序列的靶植物的实例应该包括但不限于苜蓿、苹果、杏、拟南芥、洋蓟、芝麻菜、芦笋、鳄梨、香蕉、大麦、豆类、甜菜、黑莓、蓝莓、西兰花、抱子甘蓝、卷心菜、加拿大油菜、香瓜、胡萝卜、木薯、蓖麻子、菜花、芹菜、樱桃、菊苣、芫荽、柑桔、克莱门氏小柑橘、三叶草、椰子、咖啡、玉米、棉、蔓越莓、黄瓜、花旗松、茄子、菊苣、茅菜、桉树、茴香、无花果、大蒜、葫芦、葡萄、柚子树、白兰瓜、豆薯、猕猴桃、生菜、韭葱、柠檬、酸橙、火炬松、亚麻子、芒果、甜瓜、蘑菇、油桃、坚果、燕麦、油棕、油菜、秋葵、橄榄树、洋葱、橙、观赏植物、棕榈、木瓜树、欧芹、欧洲防风草、豌豆、桃树、花生、梨树、胡椒、柿树、松树、菠萝、车前草、李树、石榴树、白杨、马铃薯、南瓜、温柏、辐射松、红菊苣、萝卜、油菜、树莓、稻、黑麦、高粱、南方松、大豆、菠菜、南瓜、草莓、甜菜、甘蔗、向日葵、甘薯、枫香树、柑橘、茶、烟草、蕃茄、黑小麦、草皮草、芜菁、葡萄树、西瓜、小麦、薯蓣和西葫芦。
组合物:
本发明的组合物是其基因组中包含本发明的任何重组dna构建体(例如上面所讨论的任何一种构建体)的植物。组合物也包括任何植物的子代,以及获取自植物或其子代的任何种子,其中所述子代或种子在其基因组中包含重组dna构建体。子代包括通过植物的自花授粉或异型杂交而获得的连续世代。子代也包括杂交种和自交系。
在杂交种子繁殖的农作物中,成熟的转基因植物可自花授粉而产生纯合的自交系植物。该自交系植物产生含有新导入的重组dna构建体的种子。这些种子可生长而产生将会表现出改变的农学特性的植物,或者可用于育种程序以产生杂交种子,这些杂交种子可生长而产生将会表现出如改变的农学特性的植物。所述种子可为玉米种子或水稻种子。
植物可为单子叶植物或双子叶植物,例如玉米或大豆植物,如玉米杂种植物或玉米自交系植物。植物还可为向日葵、高梁、油菜、小麦、苜蓿、棉花、水稻、大麦或黍。
重组dna构建体可稳定地整合进植物的基因组中。
实施方案包括但不限于以下实施方案:
1.在基因组中包含重组dna构建体的转基因植物(例如水稻、玉米或大豆植物),所述重组dna构建体包含可操作地连接至少一种异源调控序列的多核苷酸,其中所述多核苷酸编码多肽,所述多肽的氨基酸序列在与seqidno:4或7进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性,并且其中所述植物在与不包含所述重组dna构建体的对照植物进行比较时表现出改变的开花期,过量表达所述多肽促进植物营养生长向生殖生长的转变,减少该多肽的表达量延长了植物从营养生长到生殖生长的转变时间。
2.在基因组中含有重组dna构建体的转基因植物(例如水稻、玉米或大豆植物),所述重组dna构建体包含可操作连接至少一个调控序列的多核苷酸,其中所述多核苷酸编码mbd1或usp1多肽,所述植物与对照植物相比,显示改变的开花时间。当与对照植物相比,所述植物可进一步显示至少一个农艺特征的改变。
3.在基因组中含有抑制dna构建体的转基因植物(例如水稻、玉米或大豆植物),所述抑制dna构建体包含至少一个调控元件可操作连接至源于目的基因正义链或反义链的全部或部分的片段,所述片段的核苷酸序列与该片段来源的正义链或反义链全部或部分的目的基因相比,至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性,其中所述目的基因编码mbd1或usp1多肽,与对照植物相比,所述植物的开花时间延迟。
4.在基因组中含有抑制dna构建体的转基因植物(例如水稻、玉米或大豆植物),所述抑制dna构建体包含至少一个调控元件可操作连接至全部或部分(a)编码多肽的核酸序列,所述多肽的氨基酸序列与seqidno:4或7相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%得序列一致性,或者(b)(a)核酸序列的全长互补序列,其中所述植物与对照植物相比,显示延迟的开花时间。
5.实施方案1到4的转基因植物,其中多聚核苷酸编码mbd1或usp1多肽,所述mbd1或usp1多肽可来自水稻(oryzasativa)、野生稻(oryzaaustraliensis)、短舌野生稻(oryzabarthii)、非洲型稻(oryzaglaberrima)、阔叶稻(oryzalatifolia)、长雄野生稻(oryzalongistaminata)、南方野生稻(oryzameridionalis)、药用野生稻(oryzaofficinalis)、斑点野生稻(oryzapunctata)、普通野生稻(oryzarufipogon)(红稻)、印度野生稻(oryzanivara)、拟南芥(arabidopsisthaliana)、鹰嘴豆(cicerarietinum)、马铃薯(solanumtuberosum)、甘蓝(brassicaoleracea)、玉米(zeamays)、大豆(glycinemax)、烟豆(glycinetabacina)、野大豆(glycinesoja)和短绒野大豆(glycinetomentella)。
6.1-5任意实施方案的转基因植物,其中转基因植物进一步包含至少一个多核苷酸,其编码调控开花时间的多肽。
7.1-5任意实施方案的转基因植物,其中所述转基因植物进一步包含至少一个重组多核苷酸,其编码目的多肽。
8.1-7实施方案中上述植物的任意子代,实施方案1-7上述植物的任意种子,源于实施方案1-7任意上述植物的细胞和其子代。
前述实施方案1-8中的任意一项或本发明公开的其他实施方案中,重组dna构建体还包括至少一个异源的在植物中有功能的启动子作为调控序列。
下面的例子描述了一些代表性的方法和技术用于调控植物开花时间和观测和/或评估植物在该条件下的农艺特征。
1.转化的植物的子代,该转化的植物对于重组dna构建体来说是半合子的,该子代分离成包含或不包含该dna构建体的植株:包含该重组dna构建体的子代将通常相对于不包含该重组dna构建体的子代来进行测量(即,不包含该重组dna构建体的子代是对照或参照植物)。
2.重组dna构建体基因渗入至近交系中,例如在玉米中,或基因渗入进变种中,例如在大豆中:基因渗入品系将通常相对于亲本近交系或变种品系进行测量(即,亲本近交系或品种品系是对照或参照植物)。
3.双杂交系,其中第一杂交系由两个亲本近交系产生,而第二杂交系由相同的两个亲本近交系产生,不同的是其中一个亲本近交系含有重组dna构建体:第二杂交系通常将相对于第一杂交系进行测量(即第一杂交系为对照植物或参照植物)。
4.包含重组dna构建体的植物:该植株可以相对于这样的对照植物进行评价或测量,该对照植物不包含重组dna构建体,但具有与该植株相当的遗传背景(例如,与包含重组dna构建体的植株相比较,该遗传物质具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列同一性)。存在许多可用于分析、比较和表征植物遗传背景的实验室的技术;其中这些技术是同工酶电泳、限制性片段长度多态性(rflps)、随机扩增多态性dna(rapds)、任何引物聚合成酶链式反应(ap-pcr)、dna扩增指纹(daf)、序列特异扩增区域(scars)、扩增片段长度多态性
此外,本领域的普通技术人员将容易认识到,评价或测量转基因植物的农学特性或表型时合适的对照或参照植物将不包括先前已经针对所需的农学特性或表型,通过诱变或转化而选择的植物。
方法:
方法包括但不限于:调控植物开花时间的方法,观测和/或评估植物农业特征的方法,制备种子的方法。所述植物可为单子叶植物或双子叶植物,例如水稻、玉米、拟南芥、大豆植物。植物还可以是向日葵、高粱、加拿大油菜、小麦、苜蓿、棉花、大麦或黍。所述种子可为水稻、玉米、拟南芥或大豆种子,例如玉米杂交种子或玉米自交种子。
方法还包括但不限于如下方法:
转化细胞的方法包含用本发明中任意分离的多核苷酸转化细胞。也包括通过这种方法转化的细胞。在具体实施方案中,所述细胞是真核细胞,例如酵母、昆虫或植物细胞,或原核细胞例如细菌细胞。
产生转基因植物的方法,其包括用本发明的任何分离的多核苷酸或重组dna构建体来转化植物细胞并由转化的植物细胞再生转基因植物。本发明也涉及由该方法制备的转基因植物,以及从该转基因植物中获取的转基因种子。
用于从细胞或细胞培养基中分离本发明多肽的方法,其中所述细胞包含具有本发明多核苷酸的重组dna构建体,所述重组dna构建体包含本发明的多核苷酸可操作地连接到至少一种调控序列,并且其中转化的宿主细胞在适于重组dna构建体表达的条件下生长。
改变宿主细胞中本发明多肽表达水平的方法,包括:(a)用本发明的重组dna构建体转化宿主细胞;以及(b)在适于表达所述重组dna构建体的条件下使转化的细胞生长,其中重组dna构建体的表达导致转化的宿主细胞中的本发明多肽含量改变。
调控植物开花时间的方法,包括(a)将重组dna构建体导入到可再生的植物细胞中,所述重组dna构建体包含可操作地连接至少一种调控序列(例如在植物中有功能的启动子)的多核苷酸,其中所述多核苷酸编码多肽,所述所述多肽具有的氨基酸序列在与seqidno:4或7进行比较时具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;和(b)在步骤(a)之后,由所述可再生的植物细胞再生转基因植物,其中所述转基因植物在其基因组中包含所述重组dna构建体并且在与不包含所述重组dna构建体的对照植物进行比较时表现出改变的开花时间。所述方法还可包括(c)获得来源于所述转基因植物的子代植物,其中所述子代植物在其基因组中包含重组dna构建体并且在与不包含所述重组dna构建体的对照植物进行比较时表现改变的开花时间。
在一些实施方案中,控制植物开花时间的方法包括在植物中过量表达mbd1或usp1多肽,在一些实施方案中,控制开花时间的方法包括采用本发明的dna构建体转化植物或植物细胞。
生产种子的方法包括任意前述方法,还包括从所述子代植物获得种子,其中所述种子在其基因组中包含重组dna构建体。
在一些实施方案中,本发明提供的种子在其基因组中包含发明的重组dna构建体。
在前述任意的方法中或本文其他实施方案披露的任意方法中确定转基因植物农艺性状改变的步骤,如果适当,可包括确定在可变的环境条件下,与不包含重组dna构建体的对照植物相比,转基因植物是否显示至少一种农艺性状的变化。
在前述任意的方法中或本文其他实施方案披露的任意方法中确定转基因子代植物农艺性状改变的步骤,如果适当,可包括确定在可变的环境条件下,与不包含重组dna构建体的对照植物相比,转基因子代植物是否显示至少一种农艺性状的变化。
在前述任意的方法中或本文其他实施方案披露的任意方法中,所述引入步骤中所述再生植物细胞可包括愈伤细胞、胚胎愈伤细胞、配子细胞、分生细胞、或不成熟的胚胎细胞。可再生植物细胞可源于近交玉米植株。
在前述任意方法或本文披露的其他实施方案中的任意方法中,所述再生步骤可包括:(i)在含有胚促激素培养基上培养所述转化的植物细胞直至长出愈伤组织;(ii)将步骤(i)所述的转化植物细胞转移至含有组织促激素的第一培养基上;和(iii)将步骤(ii)所述转化植物细胞接种到第二培养基上,使其茎伸长、根发育或两者均发育。
在前述任意方法或本文披露的其他实施方案的任意方法中,存在将重组dna构建体导入再生植物细胞的可替代方法,所述重组dna构建体包含可操作地与至少一个调节序列连接的多核苷酸。例如,将调控序列(诸如一个或多个增强子、可作为转座元件的一部分)导入再生植物细胞中,然后筛选调控序列可操作地与即时披露的编码多肽的内源基因连接的转基因事件。
将本文披露的重组dna构建导入植物的适当的技术,包括,但不限于直接dna吸收、化学处理、电穿孔、微注射、细胞融合、转染、载体介导dna转移、轰击、或农杆菌介导的转化。植物转化和再生的技术在国际专利公开号wo2009/006276中描述,其全部内容并入作为参考。
另外,也有修饰或改变宿主内源基因组dna的方法,包括改变宿主天然dna序列或调控元件、编码或非编码序列等前体转基因序列。这些方法也可以用于将核酸序列靶向到基因组中改造目标识别序列。例如本文中转基因修饰的细胞或植物使用传统的基因工程核酸酶如产生修饰植物基因组的归位核酸内切酶产生(例如wo2009/114321;gao等(2010)plantjournal1:176-187)。其他的位点定向工程是通过使用锌指结构域识别偶联的限制性内切酶的限制性特点修饰内源基因(例如urnov等(2010)natrevgenet.11(9):636-46;shukla等(2009)nature459(7245):437-41)。转录激活因子类似-dna修饰酶(transcriptionactivator-likeeffector-dnamodifyingenzyme,tale或talen)可用于基因工程修饰植物基因组,参见例如us20110145940,cermak等(2011)nucleicacidsres.39(12)和boch等(2009),science326(5959):1509-12。植物基因组定点修饰也可以使用细菌ii型crispr(成簇的规律的间隔的短回文的重复序列,clusteredregularlyinterspacedshortpalindromicrepeats)/cas(crispr关联蛋白,crispr-associated)系统,参考例如belhaj等(2013),plantmethods9:39.crispr/cas系统可以允许可定制的小的非编码rna引导的基因组dna靶向切割。
本领域的技术人员熟悉将编码期望多肽的基因转化植物和培育再生植物的方法。再生植物可以自花授粉产生纯合子转基因植物,或者将再生植物的花粉与种子长出的农艺重要的植物杂交,或者农艺重要的植物的花粉与再生的转基因植物杂交。本领域的技术人员熟知培育本文披露的含有期望多肽的转基因植物的方法。
转基因植物中性状的堆叠
转基因植物可包含本发明公开的一个或多个花期调控多核苷酸与一个或多个另外的多核苷酸的堆叠,从而导致多个多肽序列的产生或抑制。包含多核苷酸序列的堆叠的转基因植物可通过传统育种方法或通过遗传工程方法中的一者或两者来获得。这些方法包括但不限于培育各自包含所关注多核苷酸的单独品系,用后续基因转化包含本文所公开的基因的转基因植物,以及将基因共转化进单个植物细胞中。如本文所用,术语“堆叠”包括具有存在于同一植物中的两种或更多种性状(例如,两种性状均掺入核基因组中,一种性状掺入核基因组中且一种性状掺入质体的基因组中,或者两种性状均掺入质体的基因组中)。在一个非限制性例子中,“堆叠性状”包含序列彼此物理相邻的分子堆叠。如本文所用的性状是指衍自特定序列或序列群组的表型。可使用包含多个基因的单个转化载体或单独地携带于多个载体上的基因进行基因的共转化。如果序列通过遗传转化植株来堆叠,则所关注的多核苷酸序列可在任何时间以任何顺序进行组合。可以用共转化方案将性状与所关注多核苷酸同时引入,所述多核苷酸由转化盒的任何组合提供。例如,如果将要引入两条序列,则可将该两条序列包含在单独的转化盒中(反式)或包含在同一转化盒中(顺式)。可通过相同启动子或不同启动子驱动所述序列表达。在某些情况中,可能期望引入会抑制所关注多核苷酸的表达的转化盒。这可以与其他抑制盒或过表达盒的任何组合进行组合以在植物中生成所需的性状组合。还认识到可使用位点特异性重组系统在所需的基因组位置堆叠多核苷酸序列。参见,例如,wo1999/25821、wo1999/25854、wo1999/25840、wo1999/25855和wo1999/25853,上述专利均以引用的方式并入本文。
实施例
本文的具体实施进一步在下列实例中示明。在这些实例中,除非特别说明,采用摄氏/公制。在这些实例中,仅用举例说明具体的实施过程。通过上述讨论和具体事例,本领域的专业人员可以查明本发明的基本特征,经过各种改变和修改将本发明应用于各种用途和条件,并不偏离本发明主体和范围。因此,除了本专利表述和讨论的各种修改外,本领域内的专业人员所做的不偏离本发明主题的修改也将落入本专利的权力要求范围内。
实施例1.花期调控基因的克隆和过表达载体的构建
基于基因位点loc_os05g33550.1和loc_os07g36600.1的序列信息,设计引物,并克隆水稻花期调控基因。引物序列和扩增片段的长度如表2所示。
以中花11号水稻叶、茎和根混合的cdna库为模板克隆osmbd1和osusp1基因的cdna,pcr反应混合液和pcr程序如表3和4所示。
表2.克隆水稻花期调控基因的引物
表3.pcr反应混合液
表4.克隆水稻花期调控基因的pcr循环状况
pcr扩增产物在琼脂糖凝胶电泳分离后采用柱式试剂盒回收,并与ta克隆载体连接。通过测序确定pcr产物的核酸序列和在构建体中的方向,然后将基因克隆到植物双元载体dp0158(pcambia1300-dsred,seqidno:1)中。dp0995载体中克隆的核苷酸序列和osmbd1的编码序列如seqidno:2和3所示,osmbd1的氨基酸序列如seqidno:4所示;dp0998载体中克隆的核苷酸序列和osusp1的编码序列如seqidno:5和6所示,osusp1的氨基酸序列如seqidno:7所示。
实施例2.转化获得转基因水稻
过表达载体和空载体(dp0158)采用林拥军和张启发((2005)plantcellrep.23:540-547)描述的农杆菌介导的方法转化入中花11号水稻。转化实验室获得的t0代转基因幼苗移栽至田间水田中获得t1种子,t1和t2代种子储藏在4℃冷库中。过表达载体含有dsred和hyg基因,t1和t2代种子在绿色荧光灯下发红色荧光的为转基因种子,并用于下列开花性状验证试验。
转基因水稻植株中基因表达分析:
采用标准的实时rt-pcr程序诸如源于
采集t1代灌浆期dp0995水稻植株的倒二叶,提取mrna,采用下面的引物测定osmbd1基因的表达水平。如图1所示,zh11-tc水稻中基因的表达水平设置为1.00,zh11-wt和dp0158对照水稻中基因的表达水平与zh11-tc水稻中类似,osmbd1基因在所有15个转基因株系中过量表达。
dp995-f1:5'-ctgactgctcgtgtgaagac-3'(seqidno:12)
dp995-r1:5'-tggaggctttggtatgttagg-3'(seqidno:13)
采集t1代灌浆期dp0998水稻植株的倒二叶,提取mrna,采用下面的引物测定osusp1基因的表达水平。如图2所示,zh11-tc水稻中基因的表达水平设置为1.00,zh-wt和dp0158对照水稻中基因的表达水平与zh11-tc水稻中类似,osusp1基因在所测定的14个转基因株系中过量表达,dp0998.15水稻植株中osusp1基因的表达量较低,仅为zh11-tc中基因表达量的1.87倍。
dp0998-f1:5'-ccctacaagatccacatcgtc-3'(seqidno:14)
dp0998-r1:5'-cttccaagcctccccttg-3'(seqidno:15)
实施例3.osmbd1转基因水稻开花性状的观测
t1代osmbd1转基因植株在北京田间(40°13’n,116°13’e)种植扩繁收获t2代种子,并记录植株生长过程中的表型。
方法:
t1代转基因种子于32℃800ppm多菌灵中消毒8h后,蒸馏水冲洗3~5次,然后在35-37℃培养箱中萌发36h。萌发的种子于2014年5月14日种植在田间苗床上,6月12日,三叶期时,将幼苗移栽到北京田间稻田中。每个转基因株系的10棵植株种植一行,zh11-wt(中花11号野生型)、zh11-tc(组织培养获得的中花11号)和dp0158(转化空载体dp0158的中花11号水稻)临近转基因水稻,种植在同一个小区,并用作对照。
水稻植株正常管理,并使用相应的杀虫剂和化肥,实验过程中观察并记录植株表型。
本试验记录抽穗期和成熟期,抽穗期是指一行中每棵植株50%的幼穗抽出剑叶叶鞘的日期,成熟期指每穗谷粒颖壳90%以上变黄或95%以上谷粒小穗轴及副护颖变黄的日期,此时为最佳收获时期。如果转基因水稻的抽穗期早于对照水稻植物(中花11号、zh11-tc或dp0158植株),那么转基因水稻被认为具有早抽穗的特征,转入的基因在控制植物开花时间上发挥作用。
本试验还测定了植株高度、有效穗数和单株产量。植株高度指从地表到最高穗或叶顶部的长度。收获时,将每个转基因株系的全部或位于中间位置的六株收获,先将稻穗剪下并存放在一个袋子中,然后将茎部贴地剪下放在另外的袋子中,测定每个单株的有效穗数和单株产量。植株高度、有效穗数和单株产量采用asreml混合线性模型分析。
结果:
2014年5月12日将15个osmbd1转基因水稻株系播种,于2014年6月12日移栽到北京田间。水稻植株正常管理,8月6日时,已有12个转基因株系的50%幼穗从剑叶鞘中抽出;10棵dp0995.03水稻植株中有7棵植株和10棵dp0995.07水稻植株中有8棵植株也有50%的幼穗抽出;dp0995.12水稻植株中一半在8月6日有50%有穗抽出,另一半的抽穗期在8月10日;而对照植株zh11-wt、zh11-tc和dp0158在8月21日抽穗。上述结果表明几乎所有的转基因水稻株系比对照植株早抽穗月15天。转基因水稻植株也比对照植株早成熟约15天。
表5.t1代osmbd1转基因水稻的开花性状、植株高度和产量(第一次北京验证)
抽穗期或开花期通过影响基本营养生长的时间调控作物的生物量,进而影响籽粒产量。为进一步证实这一观点,我们测定了植株高度、单株有效穗数和单株产量。表6-8表明osmbd1转基因水稻植株的单株平均籽粒产量低于zh11-wt,高于zh11-tc和dp0158两个对照,且差异均未达到显著水平;osmbd1转基因水稻植株比zh11-wt、zh11-tc和dp0158对照植株显著的矮;有效穗数与对照植株相比没有显著性差异。
表6.t1代osmbd1转基因水稻在载体水平上产量分析(第一次北京验证)
表7.t1代osmbd1转基因水稻在载体水平上植株高度分析(第一次北京验证)
表8.t1代osmbd1转基因水稻在载体水平上有效穗数分析(第一次北京验证)
收获t2代种子后,在绿色荧光灯下测定了t2代种子荧光分离比,结果表明t2代种子显示3:1的荧光和非荧光种子分离比,进一步表明单拷贝的osmbd1基因插入不同的转基因单株中。
实施例4.osmbd1转基因水稻的开花性状验证
抽穗期或开花期是一个重要的农艺性状,决定着水稻品种的适应性和地理区域分布,适当的抽穗期是获得理想产量水平的先决条件。
为进一步探讨osmbd1转基因水稻的开花性状,并探讨温度或光周期是否影响转基因水稻的抽穗期或开花期,我们将t1和t2代种子种植在不同的地理位置和环境条件下:hn(海南,18°30’n,109°3’e),bj(北京,40°13’n,116°13’e)andnx(宁夏,38°36'n,106°23'e,海拔高1106.3m)。
试验方法与实施例3描述的方法相同。
结果
t1代植株
1)2015海南
2014年11月21日,在海南田间种植了6个t1代osmbd1转基因水稻株系,zh11-tc水稻植株用作对照,12月15日将三叶期水稻移栽到稻田中。播种后84天zh11-tc对照水稻植株到达抽穗期,4个转基因株系显示出与zh11-tc相同的抽穗期,只有两个转基因株系比对照植株早抽穗3天。而早抽穗的株系与正常抽穗的四个株系成熟期没有差别,转基因水稻植株和对照植株同时成熟。
表9展示了单株平均籽粒产量,两个早抽穗株系的平均单株籽粒产量低于zh11-tc对照,正常抽穗的四个株系的单株平均产量高于zh11-tc对照。
表9.t1代osmbd1转基因水稻在海南的开花性状、植株高度和产量
2)2015北京
6个t1代osmbd1转基因株系种植在北京稻田中,zh11-tc、dp0158和dp0995.bn(t1dp0995转基因水稻种子扩繁时分离的t2代非荧光种子)用作对照。2015年4月22日将t1代转基因种子和对照种子播种,此时间早于2014年北京的播种时间。播种91天后,dp0995.15水稻植株的50%幼穗从剑叶鞘中抽出,其余5个转基因株系也显示出较早的抽穗期(表10)。这些结果进一步确认了osmbd1转基因水稻植株早抽穗/开花的性状,osmbd1转基因水稻植株比对照植株早抽穗9~14天。
如表10所示,所有6个osmbd1转基因株系均比对照植株矮;有效穗数少于zh11-tc,但与dp0158和dp0995.bn相比无差异;转基因水稻的单株平均籽粒产量少于对照植株。
表10.t1代osmbd1转基因水稻在北京的开花性状、植株高度和产量(第二次北京验证)
3)2015年宁夏
种植在北京的6个t1代osmbd1转基因株系进一步在宁夏进行验证,zh11-tc和dp0158植株用作对照。于4月14日播种育秧,5月18日移栽到田间稻田中。播种后108天,dp0995.15水稻植株的50%幼穗抽出剑叶鞘,而zh11-tc和dp0158对照水稻植株在播种后128天到达抽穗期,与zh11-tc和dp0158植株相比,osmbd1转基因水稻提前抽穗13~20天(表11)。
进一步分析表明osmbd1转基因水稻比两个对照水稻植株高,且有效穗数多余对照。osmbd1转基因水稻在九月份成熟,而由于温度较低,zh11-tc和dp0158水稻在十月初并没有完全成熟,影响了zh11-tc和dp0158的产量。osmbd1转基因水稻的平均单株籽粒产量高于zh11-tc和dp0158植株。
表11.t1代osmbd1转基因水稻在宁夏的开花性状、植株高度和产量
t2代植株
1)2015海南
将t2代7个osmbd1转基因株系的9个单株的种子种植在海南,zh11-tc水稻植株用作对照。与zh11-tc对照植株相比,dp0995.12水稻植株显示出抽穗期早7天,其余转基因株系水稻的抽穗期与zh11-tc相同。早抽穗株系的平均单株籽粒产量低于zh11-tc,而osmbd1转基因水稻的单株籽粒产量在载体水平与zh11-tc相近。
表12.t2代osmbd1转基因水稻在海南的开花性状、植株高度和产量
2)2015北京
t2代osmbd1转基因种子在北京种植两次,第一次试验的播种时间早于2014年,第二次试验的播种时间与2014年相近。两次试验中种植相同的osmbd1转基因株系,zh11-tc、dp0158和dp0995.bn水稻植株用作对照,并临近转基因水稻植株种植。
如表13和14所示,与对照相比,第一次试验中,除dp0995.03.61水稻单株外,源于6个转基因株系的11个单株的抽穗期早8~14天,第二次试验中,除dp0995.03.61和dp0995.03.62水稻单株外,10个单株的抽穗期早5~11天。这些结果进一步表明t1和t2代osmbd1转基因水稻植株在北京显示早抽穗性状。
进一步分析表明,两次试验中,osmbd1转基因水稻植株比对照植株矮;转基因水稻和对照之间有效穗数没有明显的差异;且osmbd1转基因水稻的平均单株籽粒产量低于zh11-tc对照。
表13.t2代osmbd1转基因水稻在北京的开花性状、植株高度和产量(第一次试验)
表14.t2代osmbd1转基因水稻在北京的开花性状、植株高度和产量(第二次试验)
3)2015宁夏
源于6个转基因株系的12个单株的t2代osmbd1种子种植在宁夏田间,zh11-tc和dp0158植株用作对照。播种后128天,zh11-tc和dp0158水稻植株中50%的幼穗抽出剑叶鞘,除dp0995.03.61水稻外的11个单株的抽穗期比对照早18~23天。转基因水稻植株比zh11-tc和dp0158早成熟。
植株高度和籽粒产量如表15所示,大部分osmbd1转基因水稻植株高于zh11-tc和dp0158水稻植株,且籽粒产量也高于zh11-tc和dp0158植株。zh11-tc和dp0158对照籽粒比较低的另一个原因为后期低温和抽穗期比较晚。因此早抽穗或开花能够避免成熟后期的霜降或低温造成的不良影响。
表15.t2代osmbd1转基因水稻在宁夏的开花性状、植株高度和产量
实施例5.osmbd1转基因水稻的田间干旱验证
开花期干旱胁迫是农业实践中的重要问题,osmbd转基因水稻进一步在田间干旱条件下测试。对于成熟水稻的田间干旱试验,每个构建体选择12个转基因株系,t2代种子首先按照实施例3描述的方法消毒,萌发的种子种植在田间苗床上,三叶期时,移栽到试验田地中,每个转基因株系种植一行10棵植株作为一个重复,一个小区设置四个重复,zh11-tc、dp0158和bn临近转基因株系种植,并种在一个小区中,用作统计分析的对照。
水稻植株正常管理,并施以相应的化肥和杀虫剂。水稻始穗期停止灌水,因此抽穗至开花期时能够产生干旱胁迫,干旱时间长短和胁迫程度取决于温度和湿度等天气条件。使用tdr300(spectrumtechnologies,inc.)在每块区域的十个地点每四天测定土壤体积含水量。
干旱胁迫过程中观察记录抽穗期、叶片卷曲,萎蔫,枯叶、敏旱或抗旱的表型。中午特别注意卷叶程度,收获时,每个株系的每重复挑选6株具有代表性的植株(不包括边缘植株)混合收割,脱粒并称量每份水稻材料籽粒的重量,计算出单株水稻的籽粒重量。利用asreml软件,采用混合线性模型,对单株籽粒重量进行统计分析。在p≤0.1水平下,选择阳性株系。
田间干旱试验结果:
12个osmbd1转基因株系在宁夏进行试验,zh11-tc、dp0158和dp0995.bn临近转基因株系种植,并用作对照。zh11-tc水稻植株幼穗分化ii期停止浇水至种子成熟产生干旱胁迫。如图3所示,osmbd1转基因水稻植株抽穗前,田间土壤体积含水量从45%降到10%,osmbd1转基因水稻抽穗之后和zh11-tc水稻植株抽穗后各出现了一次降雨增加了土壤含水量。与对照植株相比,大部分osmbd1转基因水稻植株早抽穗23天,dp0995.03.61、dp0995.06.61和dp0995.09.69水稻植株除外,且dp0995.03.61水稻植株比对照晚抽穗7天。早抽穗的转基因水稻比对照植株早成熟,并能在九月份完全成熟,而对照水稻植株和dp0995.03.61、dp0995.06.61和dp0995.09.69水稻植株因为后期低温不能完全成熟。osmbd1转基因水稻植株因为较早抽穗和成熟受到较短时间的干旱胁迫,因此11个转基因单株的平均单株籽粒产量均高于三个对照(表16)。上述结果表明osmbd1转基因水稻在干旱胁迫后具有较高的单株籽粒产量。
表16.osmbd1转基因水稻在田间干旱条件下的产量分析
实施例6.osusp1转基因水稻植株的开花性状观测
t1代osusp1转基因水稻种子在北京田间(40°13’n,116°13’e)扩繁用于获得t2代种子,试验过程中观察并记录表型。
试验方法如实施例3中所描述的相同。
结果:
2014年5月12日,将15个t1代osusp1转基因水稻株系的种子播种在田间的苗床上,并于6月12日移栽到北京田间的稻田中。水稻植株正常的浇水和管理,并记录植株的表型。8月4日,14个转基因株系的50%幼穗从剑叶鞘中抽出,只有一个转基因株系dp0998.15于8月16日50%的幼穗从剑叶鞘中抽出;而三个对照植株于8月21日到达抽穗期。14个转基因水稻株系的抽穗期比对照植株早17天,且osusp1基因在这14个转基因株系的叶片中高量表达;dp0998.15株系的抽穗期比对照植株早5天,osusp1基因在其叶片中的表达量相对较低。因此推测早抽穗的时间与osusp1基因在不同转基因株系中的表达量呈正相关(图2)。这些结果清楚的表明osusp1转基因水稻植株为早抽穗植物,osusp1基因可能在调控开花时间方面起重要作用。
与对照植株相比,osusp1转基因水稻植株早成熟大约19天,dp0998.15水稻株系除外。
表17.t1代osusp1转基因水稻的开花性状、植株高度和产量(第一次北京验证)
本试验还测定了植株高度,单株有效穗数和单株籽粒产量,如表18-20所示。osusp1转基因水稻植株的平均单株籽粒产量低于zh11-wt、zh11-tc和dp0158三个对照,但差别未达到显著水平;有效穗数高于zh11-wt、zh11-tc和dp0158对照植株;植株高度方面,转基因水稻植株和对照植株没有显著性差异。
表18.t1代osusp11转基因水稻在载体水平上产量分析(第一次北京验证)
表19.t1代osusp1转基因水稻在载体水平上植株高度分析(第一次北京验证)
表20.t1代osusp1转基因水稻在载体水平上有效穗数分析(第一次北京验证)
获得t2代种子后,在绿色荧光等下检测了t2代种子的荧光分离比,结果表明获得的t2代种子的荧光和非荧光分离比3:1,进一步表明单拷贝osusp1基因插入到上述不同的转基因株系中。
实施例7.osusp1转基因水稻植株的开花性状验证
最佳的产量需要抽穗期或开花期的精确调控,而抽穗期或开花期依据位置和气候而发生变化。
为进一步探讨osusp1转基因水稻的开花性状,并探讨温度或光周期是否影响转基因水稻的抽穗期或开花期,我们将t1和t2代种子种植在不同的地理位置和环境条件下:hn(海南,18°30’n,109°3’e),bj(北京,40°13’n,116°13’e)andnx(宁夏,38°36'n,106°23'e,海拔高1106.3m)。
试验方法与实施例3描述的方法相同。
结果:
t1代植株
1)2015海南
5个t1代osusp1转基因植株种植在海南田间,zh11-tc用作对照。zh11-tc对照植株播种后84天达到抽穗期,3个转基因株系的抽穗期与zh11-tc相同,1个转基因株系比对照早抽穗2天,另外一个转基因株系比对照晚抽穗2天,这5个转基因株系的成熟期与zh11-tc对照植株相同。如表21所示,所有转基因水稻株系的单株平均籽粒产量低于zh11-tc对照植株。
表21.t1代osusp1转基因水稻在海南的开花性状、植株高度和产量
2)2015北京
6个t1代osusp1转基因水稻株系种植在北京稻田中再次进行验证,zh11-tc、dp0158和dp0998.bn(t1代dp0998转基因种子扩繁时分离的t2代非荧光种子)用作对照。2015年4月22日将t1代转基因种子和对照种子播种在田间的苗床上,2015年的播种时间早于2014年。如表22所示,6个osusp1转基因株系的抽穗期均早于对照植株。这些结果进一步确认了osusp1转基因植株的早抽穗特性,且转基因植株的抽穗期比对照植株早7~14天。
osusp1转基因水稻植株也比对照植株早成熟。植株高度、有效穗数和单株产量分析表明osusp1转基因水稻植株比三个对照植株均矮,有效穗数和单株籽粒产量较少。
表22.t1代osusp1转基因水稻在北京的开花性状、植株高度和产量(第二次北京验证)
3)2015宁夏
在北京验证的6个t1代osusp1转基因株系再次在宁夏进行验证,zh11-tc和dp0158植株用作对照。与zh11-tc和dp0158对照植株相比,5个osusp1转基因株系早抽穗17~19天,dp0998.15水稻株系早抽穗8天。这些转基因植株比对照早成熟大约19天。植株高度、有效穗数和单株籽粒产量的结果表明,osusp1转基因水稻比两个对照均高,且具有较多的有效穗数。osusp1转基因水稻的单株籽粒产量高于zh11-tc和dp0158对照植株。
表23.t1代osusp1转基因水稻在宁夏的开花性状、植株高度和产量
t2代植株
1)2015海南
源于5个osusp1转基因株系的6个不同单株的t2代种子种植在海南田间,zh11-tc水稻植株用作对照。dp0998.02水稻植株比zh11-tc早抽穗3天,而其他的转基因植与zh11-tc对照植株的抽穗期相同。这些osusp1转基因水稻和zh11-tc植株成熟期相同。除dp0998.02.09和dp0998.03.02水稻植株外,osusp1转基因水稻的单株籽粒产量低于zh11-tc对照水稻植株。
表24.t2代osusp1转基因水稻在海南的开花性状、植株高度和产量
2)2015北京
t2代osusp1转基因种子在北京田间验证两次,第一次试验的播种时间早于2014年,第二次试验的播种时间与2014年相近。两次试验验证相同的osusp1转基因株系,zh11-tc、dp0158和dp0998.bn水稻植株用作对照并临近转基因株系种植。
除dp0998.15.61和dp0998.15.62外,第一次试验中osusp1转基因株系比对照早抽穗6~14天,第二次试验中早抽穗3~11天,osusp1转基因水稻植株也比对照植株早成熟。这些结果进一步表明t1和t2osusp1转基因水稻在北京田间具有早抽穗的性状。
进一步的分析显示,在两次试验中,与zh11-tc和dp0158对照植株相比,osusp1转基因水稻植株显著矮,且具有显著少的有效穗数;且osusp1转基因水稻植株的单株籽粒产量低于zh11-tc水稻植株,与dp0158和dp0998.bn水稻植株的单株籽粒产量相近。
表25.t2代osusp1转基因水稻在北京的开花性状、植株高度和产量(第一次试验)
表26.t2代osusp1转基因水稻在北京的开花性状、植株高度和产量(第二次试验)
3)2015宁夏
源于7个osusp1转基因株系的12个单株的t2代种子种植在宁夏,zh11-tc和dp0158植株用于对照。播种后128天,zh11-tc和dp0158植株到达抽穗期,而转基因水稻植株,除dp0998.15.61和dp0998.15.62外,抽穗期比对照植株早12~22天。zh11-tc和dp0158植株在播种后175天成熟,而osusp1转基因水稻会成熟的更早些,且能够避开成熟后期的低温。
植株高度和籽粒产量如表27所示,与zh11-tc和dp0158植株相比,所有osusp1转基因水稻植株较高,且籽粒产量也较高。
表27.t2代osusp1转基因水稻在宁夏的开花性状、植株高度和产量
实施例8.osusp1转基因水稻的田间干旱验证试验
osusp1转基因水稻植株在宁夏进行干旱试验,结果如下所示:
田间干旱试验结果:
12个osusp1转基因株系在宁夏进行验证,zh11-tc、dp0158和dp0998.bn临近转基因株系种植,并用作对照。zh11-tc水稻株系幼穗分化ii期停止浇水,干旱至种子成熟期产生严重干旱胁迫。osusp1转基因水稻植株抽穗前土壤体积含水量从48%降到12%,osusp1转基因水稻抽穗后和zh11-tc水稻植株抽穗后分别出现一次降雨增加了土壤体积含水量(表4)。除dp0998.15.61水稻植株外,osusp1转基因水稻植株的抽穗期比对照18~25天,dp0995.15.61株系比对照晚抽穗3天。最终,osusp1转基因水稻比对照植株早成熟,而对照植株因为后期低温不能够完全成熟。osusp1转基因水稻植株受到干旱胁迫的时间比zh11-tc和dp0158植株短。如表28所示,所有转基因事件的单株籽粒产量高于对照植株。上述结果表明osusp1水稻在干旱胁迫后,单株籽粒产量高于对照。
表28.osusp1转基因水稻在田间干旱条件下的产量分析
- 还没有人留言评论。精彩留言会获得点赞!
- 一种基因片段、载体pPlasmid‑Clearance及应用的制作方法与工艺
- 一种米曲霉脂肪酶、编码基因、载体、工程菌及其应用的制作方法与工艺
- 一种正电性的基因载体功能的聚合物荧光碳量子点的制备方法及应用与流程
- 人工优化合成的Mat基因与重组载体以及改变作物抗性的方法与流程
- 斜带石斑鱼补体C1INH基因、载体、重组菌株和蛋白及其应用的制造方法与工艺
- 一种几丁质代谢调控基因、编码蛋白、载体、工程菌及其应用的制造方法与工艺
- 用于减弱基因编辑抗病毒转移载体免疫应答的方法和组合物与流程
- 苯丙氨酸变位酶、编码基因、重组载体、宿主细胞、多重PCR引物及它们的应用的制造方法与工艺
- Hg‑snb1基因片段介导的RNAi载体及其在培育抗大豆胞囊线虫转基因植物中的应用的制造方法与工艺
- 纤维素酶及其编码基因、重组载体和应用的制造方法与工艺