一种提高鼠李糖脂产量的铜绿假单胞菌及其构建方法与流程
本发明属于生物技术领域,尤其涉及一种提高鼠李糖脂产量的铜绿假单胞菌、构建方法和利用其生产鼠李糖脂的方法。
背景技术:
鼠李糖脂是一种糖脂类生物表面活性剂,不仅具有化学表面活性剂所具有的物理化学特性,如去垢、乳化、去乳化、发泡和润湿等作用,最重要的是具有环境友好性,易降解,不会对环境造成二次污染,是目前研究最广泛的生物表面活性剂之一。在环境治理、农业生产、食品医药等领域具有广泛的潜在应用价值,但其高成本限制了它的大规模生产和应用。
目前,鼠李糖脂的生产菌株主要是铜绿假单胞菌(pseudomonasaeruginosa),主要存在鼠李糖脂的产量较低和遗传不稳定等问题。其生产性能的提高一般是通过各种理化诱变或者以质粒载体的方式过量表达某些基因,而这两种方法都会造成菌株生产性能的不稳定,前者极易发生回复突变,而后者通常还需要添加抗生素,造成环境污染和生产成本的增加,因此迫切需要新的构建鼠李糖脂高产菌株的方法。
鼠李糖脂的代谢途径比较复杂,在铜绿假单胞菌中,鼠李糖脂的合成依赖于三条代谢途径,分别为脂质前体β-羟基脂肪酸(haas)合成途径、糖基前体dtdp-l-鼠李糖合成途径以及将脂质前体和糖基前体进行聚合形成单鼠李糖脂和双鼠李糖脂。鼠李糖脂合成的中的糖基前体dtdp-l-鼠李糖主要来自于ed途径,d-葡萄糖-6-磷酸在葡萄糖磷酸变位酶的作用下生成1-磷酸-葡萄糖,然后在rmlabcd的作用下生成dtdp-l-鼠李糖。dtdp-l-鼠李糖除了作为鼠李糖脂合成的前体外,同时也是胞外多糖psl和藻酸盐合成的前体,铜绿假单胞菌还会分泌另一种富含葡萄糖残基的胞外多糖pel。脂质前体β-羟基脂肪酸(haas)一部分来自于脂肪酸的重头合成途径,另一部分来自于脂肪酸的β-氧化途径。羟基脂肪酸除了参与鼠李糖脂的合成外,还参与聚羟基脂肪酸pha的合成。另外,在铜绿假单胞菌中,鼠李糖脂的合成途径还受到与群集效应相关的转录调控基因和全局性调控因子的控制。这些调控因子与鼠李糖脂代谢途径紧密关联,同时外界环境条件,如温度、通气量、碳氮源以及微量元素,尤其是铁都与鼠李糖脂的合成密切相关。正是如此,在现有技术基础上,要想进一步提高鼠李糖脂具有很大困难。适宜生产鼠李糖脂的生产菌株在构建时,不仅需要考虑鼠李糖脂的产量问题,而且还需要综合考虑是否稳定遗传以及下游产品的分离纯化等问题。因此,获得同时具有鼠李糖脂的产量高、可稳定遗传以及有利于下游产品的分离纯化的鼠李糖脂生产菌是很困难的事情。
技术实现要素:
为解决上述技术问题,本发明提供一种提高鼠李糖脂产量的铜绿假单胞菌及其构建方法,该重组铜绿假单胞菌菌株具有显著提高鼠李糖脂的产量、稳定遗传以及有利于下游产品的分离纯化等优点。利用该重组铜绿假单胞菌生产鼠李糖脂,重组铜绿假单胞菌与作为出发菌的铜绿假单胞菌相比,鼠李糖脂合成能力提高,发酵液澄清不粘稠,发酵液中胞外多糖量减少,后续处理容易,鼠李糖脂纯度提高。
本发明解决上述技术问题的技术方案如下:
本发明提供一种提高鼠李糖脂产量的铜绿假单胞菌,该铜绿假单胞菌的a组中的基因表达量降低或丧失,同时,该铜绿假单胞菌的b组中的基因表达量升高;
a组中的基因选自pslabcdefghijklmno胞外多糖合成基因簇、pelabcdefg多糖合成基因簇、藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf以及聚羟基脂肪酸pha合成关键基因簇phac1-d-c2中的一个基因或任意几个基因的组合;
b组中的基因选自rmlacbd、rmlbdac、rhlyz、algc、fadd4、lipc以及esta中的一个基因或任意几个基因的组合。
本发明的有益效果是:
a组中的基因主要与鼠李糖脂前体合成的旁路相关,敲除该组基因将有利于减少前体的消耗,b组中的基因为直接参与鼠李糖脂前体合成途径中的相关基因,过量表达该组基因,将有利于增加鼠李糖脂前体的合成量。在研究中发现,将a组中的基因与b组中的基因配合操作有利于提高鼠李糖脂的产量。
本发明通过使这些与鼠李糖脂前体合成的旁路相关的基因表达量降低或丧失,有助于增加鼠李糖脂的产量,更进一步的,将过量表达的基因如rmlacbd(或rmlbdac)、rhlyz、algc、fadd4、lipc以及esta整合到染色体上,不仅能够增加铜绿假单胞菌的油脂利用能力,而且可以使这些基因得到稳定遗传。
进一步,a组中的基因包括pslabcdefghijklmno胞外多糖合成基因簇、pelabcdefg多糖合成基因簇、藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf以及聚羟基脂肪酸pha合成关键基因簇phac1-d-c2;b组中基因包括rmlacbd、rhlyz、algc、fadd4、lipc以及esta或b组中的基因包括rmlbdac、rhlyz、algc、fadd4、lipc以及esta;
该铜绿假单胞菌的基因组的a组中的基因的表达量均降低或丧失,同时,该铜绿假单胞菌的基因组的b组中的基因的表达量均升高。
即该铜绿假单胞菌的基因组的pslabcdefghijklmno胞外多糖合成基因簇、pelabcdefg多糖合成基因簇、藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf以及聚羟基脂肪酸pha合成关键基因簇phac1-d-c2的表达量均降低或丧失,同时,该铜绿假单胞菌的基因组的rmlacbd、rhlyz、algc、fadd4、lipc以及esta的基因表达量均升高。
采取上述方案的有益效果为:
本发明提供的铜绿假单胞菌在铜绿假单胞菌pao1菌株中连续敲除胞外多糖合成基因簇psla-o共15个基因和pela-g共7个基因、藻酸盐合成基因簇algd-f共11个基因以及聚羟基脂肪酸合成关键基因phac1-d-c2,且在敲除基因的位置上分别插入过量表达的基因rmlacbd(或rmlbdac)、rhlyz-algc、fadd4-lipc及esta。
本发明利用基因工程技术来改造其基因组,通过敲除或者插入相关基因,减少鼠李糖脂合成前体的dtdp-l-鼠李糖和β-羟基脂肪酸(haas)的消耗,同时通过增加脂肪酶lipc的表达增加油脂的降解,促进脂肪酸β-氧化;增加rmlacbd的表达,促进dtdp-l-鼠李糖的合成,从而为鼠李糖脂的合成提供更多的脂质前体和糖基前体,最终达到提高鼠李糖脂产量的目的,同时由于敲除胞外多糖合成基因簇,使胞外多糖产量下降,更有利于下游产品的分离纯化。本发明选择在基因组上进行操作,并非以携带质粒的形式,将过量表达的基因如rmlacbd(或rmlbdac)、rhlyz、algc、fadd4、lipc以及esta整合到染色体上,不仅能够增加铜绿假单胞菌的油脂利用能力,而且可以使这些基因得到稳定遗传。
本发明获得的基因工程菌具有显著提高鼠李糖脂的产量、稳定遗传以及有利于下游产品的分离纯化等优点,适合用于鼠李糖脂的生产。实验结果表明,与作为出发菌株的铜绿假单胞菌相比,本发明提供的重组铜绿假单胞菌的鼠李糖脂产量为43.7g/l,较出发菌株提高了93.4%,而且发酵液不粘稠,易处理。
进一步,该铜绿假单胞菌的基因组中,在pslabcdefghijklmno胞外多糖合成基因簇的缺失位置处过表达rmlacbd,在pelabcdefg多糖合成基因簇的缺失位置处过表达rhlyz和algc,在藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf的缺失的位置处过表达fadd4和lipc,在聚羟基脂肪酸pha合成关键基因簇phac1-d-c2缺失的位置处过表达esta。
采取上述方案的有益效果为:本发明提供的铜绿假单胞菌具有显著提高鼠李糖脂的产量、稳定遗传以及有利于下游产品的分离纯化等优点,适合用于鼠李糖脂的生产。
本发明还提供一种上述铜绿假单胞菌的构建方法,在铜绿假单胞菌中将a组中的基因敲除或敲降,同时,在铜绿假单胞菌中将b组中的基因过表达。
进一步,所述a组基因包括pslabcdefghijklmno胞外多糖合成基因簇、pelabcdefg多糖合成基因簇、藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf以及聚羟基脂肪酸pha合成关键基因簇phac1-d-c2;所述b组基因包括rmlacbd、rhlyz、algc、fadd4、lipc以及esta;提高鼠李糖脂产量的铜绿假单胞菌的构建方法包括以下步骤:
1)以铜绿假单胞菌pao1为出发菌株,将出发菌株的pslabcdefghijklmno胞外多糖合成基因簇敲除的同时将dtdp-l-鼠李糖前体合成的关键基因rmlacbd基因簇整合到psl缺失的位置,rmlacbd基因簇受lac启动子调控,得到菌株pa111;
2)在菌株pa111的基础上,连续敲除pelabcdefg多糖合成基因簇,同时将受lac启动子调控的烯酯酰-coa水合酶/异构酶rhlyz和d-葡萄糖-6-磷酸在葡萄糖磷酸变位酶algc整合到pel基因簇缺失的位置,得到菌株pa112;
3)在菌株pa112的基础上,连续敲除藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf,同时将受lac启动子调控的脂肪酸激活蛋白fadd4和脂酶lipc整合到alg基因缺失的位置,得到菌株pa113;
4)在菌株pa113的基础上,连续敲除聚羟基脂肪酸pha合成关键基因簇phac1-d-c2,同时将受lac启动子调控的酯酶esta整合到pha基因簇敲除的位置,得到菌株pa114。
采取上述方案的有益效果为:
本发明提供的铜绿假单胞菌,首先该菌株的构建包括:1)通过敲除铜绿假单胞菌pao1鼠李糖脂合成途径中的旁路,包括敲除胞外多糖合成基因簇psla-o共15个基因和pela-g共7个基因、藻酸盐合成基因簇algd-f共11个基因以及聚羟基脂肪酸合成关键基因phac1-d-c2;2)在敲除的同时,在敲除基因的位置上插入过量表达的基因,过量表达的基因包括rmlacbd、rhlyz、algc、fadd4、lipc以及esta;3)发酵所述重组铜绿假单胞菌,得到鼠李糖脂。实验结果表明,与作为出发菌的铜绿假单胞菌pao1相比,重组铜绿假单胞菌的鼠李糖脂产量为43.7g/l,较出发菌株提高了93.4%,而且发酵液不粘稠,易处理。
本发明还提供一种利用上述铜绿假单胞菌生产鼠李糖脂的方法,包括以下步骤:将上述铜绿假单胞菌发酵,得到鼠李糖脂。
采取上述方案的有益效果为:本发明采用上述方法生产鼠李糖脂具有显著提高鼠李糖脂的产量、稳定遗传以及有利于下游产品的分离纯化等优点。
附图说明
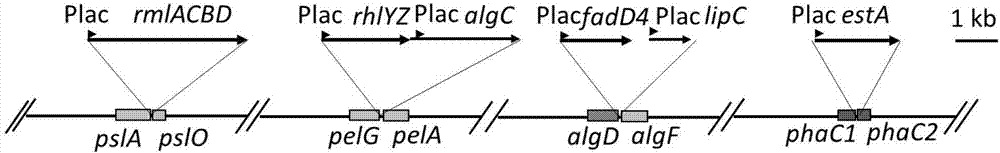
图1为psl、pel操纵子、藻酸盐和pha合成的关键基因簇示意图。
图2为敲除和敲入基因簇在铜绿假单胞菌染色体上的位置示意图。
图3为分别用苔黑酚法和称重法测定菌株pao1、菌株pa111、菌株pa112、菌株pa113和菌株pa114合成的鼠李糖脂浓度图。
图4为菌株pao1、菌株pa111、菌株pa112、菌株pa113和菌株pa114鼠李糖脂纯度图。
图5为出发菌株pao1与重组菌株pa114发酵液及提纯的鼠李糖脂比较结果。
具体实施方式
以下结合附图对本发明的原理和特征进行描述,所举实例只用于解释本发明,并非用于限定本发明的范围。
本发明通过敲除铜绿假单胞菌pao1鼠李糖脂合成途径中的旁路以及从而将过量表达的基因整合到染色体上,达到稳定遗传和提高鼠李糖脂产量的目的。需要敲除的基因有:胞外多糖合成基因簇psla-o共15个基因和pela-g共7个基因、藻酸盐合成基因簇algd-f共11个基因以及聚羟基脂肪酸合成关键基因phac1-d-c2(图1);在敲除基因的位置上同时插入需要过量表达的基因,需要过量表达的基因包括rmlacbd、rhlyz、algc、fadd4、lipc以及esta(图2)。
构建上述具有稳定遗传特性的重组铜绿假单胞菌的方法,包括以下步骤:
1)为了便于对转基因菌株进行鉴定及筛选,对用于过表达的载体pucp18和用于构建突变菌株的自杀性质粒pk18mobsacb进行加工,插入编码庆大霉素抗性基因的抗性盒,以便于利用庆大霉素抗性对转化子进行筛选。
2)筛选对鼠李糖脂提高有帮助的基因用于作为插入的过量表达基因。本发明中共过量表达了rmlacbd及其不同组合rmlbdac、algc、lipc、liph、fadd1、fadd2、fadd4、rhlyz、glpfk、rhlab等基因或基因簇,其中rmlbdac、rmlacbd、algc、liph、lipc、rhlyz单独过量表达时鼠李糖脂产量有明显提高,rmlacbd排列方式较rmlbdac更好,单独过量表达lipc的效果优于liph,fadd1、fadd2过量表达时鼠李糖脂产量反而降低,fadd4、glpfk、rhlab过量表达对鼠李糖脂的产量影响不明显;由于fadd基因为脂肪酸激活蛋白,并在铜绿假单胞菌中有多个拷贝,这些fadd基因对于不同长度脂肪酸的激活作用不同,因此在过量表达时对鼠李糖脂产量的影响也不同。在本研究中进一步选取rmlacbd、algc、lipc、fadd4、rhlyz及esta等6个基因或基因簇作为过量表达的基因。
3)构建具有遗传稳定性的高产鼠李糖脂铜绿假单胞菌。
a)以铜绿假单胞菌pao1为出发菌株,将出发菌株的基因组的pslabcdefghijklmno多糖合成基因簇敲除的同时将dtdp-l-鼠李糖前体合成的关键基因rmlacbd基因簇整合到psl缺失的位置,rmlabcd基因簇受lac启动子调控,得到菌株pa111;
b)在菌株pa111的基础上,连续敲除pelabcdefg多糖合成基因簇,同时将受lac启动子调控的烯酯酰-coa水合酶/异构酶rhlyz和d-葡萄糖-6-磷酸在葡萄糖磷酸变位酶algc整合到pel基因簇缺失的位置,菌株pa112;
c)在菌株pa112的基础上,连续敲除藻酸盐合成基因簇algd-alg8-alg44-algkegxlijf,同时将受lac启动子调控的脂肪酸激活蛋白fadd4和脂酶lipc整合到alg基因缺失的位置,菌株pa113;
d)在菌株pa113的基础上,连续敲除聚羟基脂肪酸pha合成关键基因簇phac1-d-c2,同时将受lac启动子调控的酯酶esta整合到pha基因簇敲除的位置,得到菌株pa114。
在上述的构建方法中,基因敲除顺序可以改变,过量表达基因的插入位点也可以是敲除基因簇中的任意一个位置,过量表达基因也可以变更组合,所得重组菌株应与上述菌株具有同样的功能。
提高铜绿假单胞菌鼠李糖脂合成能力的方法,包括构建具有遗传稳定性的铜绿假单胞菌的步骤,所述重组铜绿假单胞菌的鼠李糖脂合成能力高于所述作为出发菌株的铜绿假单胞菌pao1,
1)在出发菌株铜绿假单胞菌pao1的基础上,构建pao1(δpsl::rmlabcd)重组菌株,命名为菌株pa111,与出发菌株相比,鼠李糖脂产量显著提高;
2)在步骤1)得到的菌株pa111基础上,构建pa111(δpel::rhlyz-algc)重组菌株,命名为菌株pa112,与pa111菌株相比,鼠李糖脂产量提高;
3)在步骤2)得到的菌株pa112的基础上,构建pa112(δalg::fadd4-lipc)重组菌株,命名为菌株pa113,与pa112菌株相比,鼠李糖脂产量提高;
4)在步骤3)得到的菌株pa113的基础上,构建pa113(δpha::esta)重组菌株,命名为菌株pa114,与pa113菌株相比,鼠李糖脂产量提高。
制备鼠李糖脂的方法,包括下面的步骤:
1)构建与作为出发菌株的铜绿假单胞菌相比,鼠李糖脂合成前体的旁路敲除菌株以及鼠李糖脂合成前体的基因过量表达菌株;
2)发酵所述重组铜绿假单胞菌,得到鼠李糖脂。
下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。
若无特别说明,本发明所采用的专业术语与本领域技术人员所熟悉的意义相同。
下述实施例中所用材料、试剂等,如无特殊说明,均可从商业途径得到。
铜绿假单胞菌pao1(zhuk,rockco.rhlaconvertsb-hydroxyacyl-acylcarrierproteinintermediatesinfattyacidsynthesistotheb-hydroxydecanoyl-b-hydroxydecanoatecomponentofrhamnolipidsinpseudomonasaeruginosa.jbacteriol.2008,5;190(9):3147-3154.)公众可从中国科学院微生物研究所获得,该生物材料只为重复本发明的相关实验所用,不可作为其它用途使用。
下述实施例中的pk18mobsacb自杀性载体(schafera..taucha..smallmobilizablemulti-purposecloningvectorsderivedfromtheescherichiacoliplasmidspk18andpk19:selectionofdefineddeletionsinthechromosomeofcorynebacteriumglutumicum.gene.1994;145(1):69-73。)、质粒pps856(choikh,schweizerhp.animprovedmethodforrapidgenerationofunmarkedpseudomonasaeruginosadeletionmutants.bmcmicrobiol.2005,5(30):1-11)以及pucp18载体(schweizerhp.escherichia-pseudomonasshuttlevectorsderivedfrompuc18/19.gene.97(1):109-112.)公众可从中国科学院微生物研究所获得,该生物材料只为重复本发明的相关实验所用,不可作为其它用途使用。
实施例1、具有庆大霉素(gm)抗性的高拷贝质粒载体pucp18的构建
通过分析高拷贝质粒载体pucp18和庆大霉素抗性盒的序列设计引物,用于扩增庆大霉素抗性的引物分别为:
gm(ndei)-f:gtgccatatgaagttcctatactttc(seqidno.1)
gm(ndei)-r:cgcttactcatatggaattctagatc(seqidno.2)
从pps856质粒为模板扩增gm抗性基因,然后用ndei进行酶切,克隆到用ndeⅰ酶切并去磷酸化的pucp18载体中,得到具有庆大霉素(gm)抗性的质粒载体pucp18gm。
实施例2、rmlacbd和rmlbdac过表达菌株pao1/rmlacbd、pao1/rmlbdac和对比菌株pao1/pucp18gm的构建及其鼠李糖脂合成量的测定。
铜绿假单胞菌中rmlbdac操纵子编码dtdp-l-鼠李糖合成的酶,在染色体上其基因排列顺序为rmlbdac,而且rmlb基因前的启动子受群集效应和σs的调控,但在dtdp-l-鼠李糖的合成中,酶的作用顺序为rmla、rmlb、rmlc和rmld,rmla为负责将胸腺核苷酸转移到葡萄糖-1-磷酸上,生成dtdp-1-葡萄糖,为该系列反应的限速酶。rmlabcd是以二聚体的二聚体形式在起催化作用,因此我们同时构建了以rmlacbd的排列顺序置于组成型启动子之下,比较这两种排布方式对鼠李糖脂产量的影响。
首先,将rmlbdac基因克隆到pucp18gm载体中。以pao1染色体dna为模板,以prmlb-f和prmlc-r为引物,扩增rmlbdac基因片段,用saci+hindiii酶切后克隆到同样用saci+hindiii酶切的pucp18gm载体中。得到prmlbdac-pucp18gm质粒。
其次,将rmlacbd基因克隆到pucp18gm载体中。以铜绿假单胞菌pao1染色体dna为模板,分别以prmlac-f和prmlacbd-r、prmlacbd-f和prmlbd-r为引物分别扩增rmlac片段和rmlbd片段,然后通过overlappcr,以回收片段为模板,以prmlac-f和prmlbd-r为引物进行基因融合,得到rmlacbd片段,将该片段用saci+hindiii进行双酶切,克隆到同样用saci+hindiii进行双酶切的pucp18gm载体中,得到prmlacbd-pucp18gm质粒。
最后,构建pao1/rmlacbd、pao1/rmlbdac过表达菌株和对比菌株pao1/pucp18gm。分别提取pucp18gm、prmlbdac-pucp18gm和prmlacbd-pucp18gm质粒,通过电击转化,将上述三个质粒分别转入到铜绿假单胞菌pao1中,得到过表达菌株pao1/rmlacbd、pao1/rmlbdac和对比菌株pao1/pucp18gm。pcr验证,确定正确的质粒已经转入到铜绿假单胞菌中。分别测定过表达菌株pao1/rmlacbd、pao1/rmlbdac和对比菌株pao1/pucp18gm的鼠李糖脂产量。首先将pao1/rmlacbd、pao1/rmlbdac过表达菌株和对比菌株pao1/pucp18gm分别接种到lb营养肉汤中培养16-24h,然后转接到以棕榈油为碳源的ms培养基中,200rmp,37℃条件下培养96h,然后利用苔黑酚法测定鼠李糖脂的产量。结果表明过量表达rmlacbd和rmlbdac能够提高鼠李糖脂的产量22%和13%,优选的,过量表达rmlacbd效果更好。
prmlb-f:tatgagctcaatgacgattctcgtgac(seqidno.3)
prmlc-r:gaataagctttcaagacgaacgaacctg(seqidno.4)
prmlac-f:cagggagctctatgaaacgcaagggcatcatc(seqidno.5)
prmlacbd-r:gagtcataccttgttcaagcagtgtcaggggaagcagtcggcgtc(seqidno.6)
prmlacbd-f:gacgccgactgcttcccctgacactgcttgaacaaggtatgactc(seqidno.7)
prmlbd-r:atcaagcttcatcatagtggtccctgctcgctcag(seqidno.8)
实施例3、rhlyz和algc过表达菌株pao1/algc、pao1/rhlyz的构建及其鼠李糖脂合成量的测定
rhlyz为烯脂酰-coa水合酶/异构酶,是最近在铜绿假单胞菌中发现的具有脂肪酸降解转轨作用的一组酶,负责将脂肪酸β-氧化中间产物直接转化为羟脂酰-coa(abdel-mawgoud,2014)。algc蛋白同时具有磷酸葡萄糖变位酶和磷酸甘露糖变位酶两种功能,可以将葡萄糖-6-磷酸和甘露糖-6-磷酸分别转化为葡萄糖-1-磷酸和甘露糖-1-磷酸,葡萄糖-1-磷酸在rmlabcd的作用下生成dtdp-l-鼠李糖。因此推测rhlyz和algc的表达对鼠李糖脂的合成有影响。
首先,分别将rhlyz和algc基因克隆到pucp18gm载体中。以铜绿假单胞菌pao1染色体dna为模板,分别以prhlyz-f和prhlyz-r、algc-f和algc-r为引物扩增rhlyz和algc基因,rhlyz基因片段用saci+bamhi酶切后克隆到用saci+bamhi酶切的pucp18gm载体中,algc基因片段用bamhi+hindiii进行双酶切,克隆到同样用bamhi+hindiii进行双酶切的pucp18gm载体中,分别得到palgc-pucp18gm质粒和prhlyz-pucp18gm质粒。
prhlyz-f:aagagctccatgaacactgccgtcgaac(seqidno.9)
prhlyz-r:aaggatccagcttagacgctatggacgtg(seqidno.10)
algc-f:cgggatccatgagcactgcaaaagcaccgac(seqidno.11)
algc-r:ccaagcttacatgtcagaagggcacgggca(seqidno.12)
然后,构建pao1/algc、pao1/rhlyz过表达菌株。分别提取palgc-pucp18gm和prhlyz-pucp18gm质粒,通过电击转化,将这两个质粒分别转入到铜绿假单胞菌pao1中,得到pao1/algc、pao1/rhlyz过表达菌株。pcr验证,确定正确的质粒已经转入到铜绿假单胞菌中。分别测定pao1/algc、pao1/rhlyz过表达菌株和对比菌株pao1/pucp18gm的鼠李糖脂产量,结果表明过量表达rhlyz和algc能够提高鼠李糖脂的产量53.8%和32.5%。
实施例4、lipc和liph过表达菌株pao1/lipc、pao1/liph的构建及其鼠李糖脂产量的测定
铜绿假单胞菌会分泌多种蛋白酶和脂肪酶,脂肪酶lipc的缺失会显著减少胞外鼠李糖脂的合成量(frankrosenau,femsmicrobiollett309(2010),25-34)。以油脂为碳源时,油脂降解为脂肪酸和甘油是油脂利用的第一步,因此也是鼠李糖脂合成中的关键限速步骤之一,过量表达脂肪酶lipc及其折叠酶liph将有助于提高油脂的利用效率。
首先,分别将lipc和liph基因克隆到pucp18gm载体中。以铜绿假单胞菌pao1染色体dna为模板,分别以plipc-f和plipc-r、pliph-f和pliph-r为引物扩增lipc和liph基因,含lipc基因片段的pcr产物用kpni+psti酶切后克隆到用kpni+psti酶切的pucp18gm载体中,liph基因片段用saci+bamhi酶切后克隆到用saci+bamhi酶切的pucp18gm载体中,分别得到plipc-pucp18gm质粒和pliph-pucp18gm质粒。
plipc-f:ataaggtaccgatgaacaagaacaagac(seqidno.13)
plipc-r:atctgcaggttcatatccatcgtg(seqidno.14)
pliph-f:attagagctcggtgccggcccctcag(seqidno.15)
pliph-r:ccaggatcctgagggtagcgttatc(seqidno.16)
然后,构建pao1/lipc、pao1/liph过表达菌株。分别提取plipc-pucp18gm和pliph-pucp18gm质粒,通过电击转化,将这两个质粒分别转入到铜绿假单胞菌pao1中,得到pao1/lipc、pao1/liph过表达菌株。pcr验证,确定正确的质粒已经转入到铜绿假单胞菌中。分别测定pao1/lipc、pao1/liph过表达菌株和对比菌株pao1/pucp18gm的鼠李糖脂产量,结果表明过量表达lipc和liph能够提高鼠李糖脂的产量26.1%和32.5%。
实施例5、fadd1、fadd2和fadd4过表达菌株pao1/fadd1、pao1/fadd2和pao1/fadd4的构建及其鼠李糖脂合成量的测定
在假单胞菌中存在着至少6个脂酰-coa合成酶基因(fadd),其中fadd1能够被c18:1的脂肪酸强烈诱导表达,而fadd2能够被c8:0和c10:0的脂肪酸强烈诱导,而fadd4对不同碳链长度的脂肪酸均有激活作用。发明人推测过量表达fadd1、fadd2和fadd4能够增加脂肪酸的降解,从而影响鼠李糖脂的产量。
首先,分别将fadd1、fadd2和fadd4克隆到pucp18gm载体中。以铜绿假单胞菌pao1染色体dna为模板,分别以pfadd1-f和pfadd1-r、pfadd2-f和pfadd2-r、pfadd4-f和pfadd4-r为引物分别扩增fadd1、fadd2和fadd4基因,含fadd1、fadd2基因片段的pcr产物用kpni+hindiii酶切并克隆到用相同酶切的pucp18gm载体中,含fadd4基因片段的pcr产物用kpni+psti酶切后克隆到用kpni+psti酶切的pucp18gm载体中,分别得到pfadd1-pucp18gm、pfadd2-pucp18gm质粒和pfadd4-pucp18gm质粒。
pfadd1-f:ggggtaccaatgatcgaaaacttctggaagg(seqidno.17)
pfadd1-r:cccaagcttttacttctggcccgctttcttc(seqidno.18)
pfadd2-f:ggggtaccaatgcaacctgaattctgg(seqidno.19)
pfadd2-r:cccaagcttcgtttcagagccttgtaag(seqidno.20)
pfadd4-f:attagagctccgtggtcactgcaaatcgtctg(seqidno.21)
pfadd4-r:atatggatccagaggctcatgggaattccttg(seqidno.22)
然后,构建fadd1、fadd2和fadd4过表达菌株pao1/fadd1、pao1/fadd2和pao1/fadd4过表达菌株。分别提取fadd1-pucp18gm、fadd2-pucp18gm和fadd4-pucp18gm质粒,通过电击转化,将这三个质粒分别转入到铜绿假单胞菌pao1中,得到pao1/fadd1、pao1/fadd2和pao1/fadd4过表达菌株。pcr验证,确定正确的质粒已经转入到铜绿假单胞菌中。分别测定pao1/fadd1、pao1/fadd2和pao1/fadd4过表达菌株和对比菌株pao1/pucp18gm的鼠李糖脂产量,结果出乎意料的是,过量表达fadd1和fadd2使鼠李糖脂的产量分别降低了50%和69%,而fadd4对鼠李糖脂的产量基本无影响。
实施例6、glpfk和rhlab过表达菌株pao1/glpfk和pao1/rhlab的构建及其鼠李糖脂合成量的测定
glpk为甘油激酶,是甘油代谢中的限速酶,glpf为促进甘油吸收的蛋白。当以油脂为碳源时,油脂降解为脂肪酸和甘油,甘油进入微生物体内后,在磷酸甘油激酶的催化下,先形成磷酸甘油,然后再磷酸甘油脱氢酶的作用下,生成磷酸二羟丙酮,能够为鼠李糖脂的合成提供糖基前体。在铜绿假单胞菌中,鼠李糖脂的合成主要包括三个连续的反应,首先是由rhla催化两个β-羟基酯酰coa和/或acp形成β-羟基脂肪酸二聚体(haa),然后由鼠李糖基转移酶irhlb催化haa和dtdp-l-鼠李糖反应生成单鼠李糖脂,最后由鼠李糖基转移酶iirhlc催化单鼠李糖脂和dtdp-l-鼠李糖反应生成双鼠李糖脂。因此rhlab基因是鼠李糖脂合成的关键基因。为了研究glpfk和rhlab过表达对鼠李糖脂产量的影响,我们也构建了glpfk和rhlab过表达菌株pao1/glpfk和pao1/rhlab,并测定其鼠李糖脂合成量。
首先,分别将glpfk和rhlab操纵子克隆到pucp18gm载体中,以pao1染色体dna为模板,分别以pglpf-f和pglpk-r、prhlab-f和prhlab-r为引物分别扩增glpfk和rhlab基因,含glpfk基因片段的pcr产物用saci+hindiii酶切后克隆到用同样酶切的pucp18gm载体中,含rhlab基因片段的pcr产物用kpni+hindiii酶切并克隆到用相同酶切的pucp18gm载体中,分别得到pglpfk-pucp18gm质粒和prhlab-pucp18gm质粒。
pglpf-f:acgtgagctctacaagaacagacaagagag(seqidno.23)
pglpk-r:aataagcttgcaaacgttacccagggcat(seqidno.24)
prhlab-f:ggggtaccaatgcggcgcgaaagtctgtt(seqidno.25)
prhlab-r:cccaagcttaccgttcaggacgcagccttca(seqidno.26)
然后,构建glpfk和rhlab过表达菌株pao1/glpfk和pao1/rhlab过表达菌株。分别提取pglpfk-pucp18gm和prhlab-pucp18gm质粒,通过电击转化,将这两个质粒分别转入到铜绿假单胞菌pao1中,得到pao1/glpfk和pao1/rhlab过表达菌株。pcr验证,确定正确的质粒已经转入到铜绿假单胞菌中。分别测定pao1/glpfk和pao1/rhlab过表达菌株和对比菌株pao1/pucp18gm的鼠李糖脂产量,结果出乎意料的是,过量表达glpfk和rhlab对鼠李糖脂的产量都没有明显的影响。
实施例7、铜绿假单胞菌pa111、pa112、pa113和pa114菌株的构建
1)以铜绿假单胞菌pao1作为出发菌株,首先构建pa111重组菌株。构建过程如下:
(1)构建适合于在铜绿假单胞菌中筛选用的自杀性质粒载体pk18gm。以pk18-gm(bamhi)-f和pk18-gm(ncoi)-r为引物,从pps856质粒为模板扩增gm抗性基因,然后用bamhi+ncoi进行双酶切,克隆到用bglⅱ+ncoⅰ双酶切的pk18mobsacb载体中,得到自杀性质粒pk18gm。
(2)构建自杀性质粒pk18-psl-rmlacbd。以铜绿假单胞菌pao1为模板,分别以ppslup-f和ppslup-r、ppsldown-f和ppsldown-r扩增psla上游序列和pslo下游序列,用bamhi+xhoi进行双酶切,再克隆到用bamhi+hindiii酶切的pk18gm中,得到pk18-psl质粒。将pk18-psl质粒用xhoi进行酶切,回收,同时以实施例2中构建的prmlacbd-pucp18gm质粒为模板,以psl-rmlacbd-f和psl-rmlacbd-r为引物扩增rmlacbd片段,回收,通过无缝克隆拼接试剂盒(theseamlesscloningandassemblykit,thermoscientificco.)进行连接,转化e.colidh5α,得到自杀性质粒pk18-psl-rmlacbd。
(3)利用三亲结合的方法(受体菌株为pao1,供体菌为pk18-psl-rmlacbd,助质粒为prk2013)将自杀性质粒转入到pao1,通过同源重组双交换,在敲除psl基因簇的同时将rmlacbd基因簇整合到染色体上,获得pa111重组菌株。方法参照文献schafera,taucha.;smallmobilizablemulti-purposecloningvectorsderivedfromtheescherichiacoliplasmidspk18andpk19:selectionofdefineddeletionsinthechromosomeofcorynebacteriumglutumicum.gene,145(1994)69-73。
pk18-gm-f:gctgaaggatcctatactttctagag(seqidno.27)
pk18-gm-r:cttactccatggaattctagatcg(seqidno.28)
ppslup-f:ataggatccctgccgccgcgctactac(seqidno.29)
ppslup-r:agactcgagtggatagggaagagcaag(seqidno.30)
ppsldown-f:atactcgagtttgctggctttcatgaac(seqidno.31)
ppsldown-r:attaagcttggagagatcagaaacgggcgaac(seqidno.32)
psl-rmlacbd-f:cttgctcttccctatccactcagctggcacgacaggtttc(seqidno.33)
psl-rmlacbd-r:gttcatgaaagccagcaaaccatcatagtggtccctgctcgctc(seqidno.34)
2)pa112重组菌株的构建。构建过程如下:
(1)构建自杀性质粒pk18-pel-rhlyz-algc。以铜绿假单胞菌pao1为模板,分别以ppelup-f和ppelup-r、ppeldown-f和ppeldown-r扩增pela上游序列和pelg下游序列,用bamhi+xhoi进行双酶切,再克隆到用bamhi+hindiii酶切的pk18gm中,得到pk18-pel质粒。将pk18-pel质粒用xhoi进行酶切,同时以实施例3中构建的prhlyz-pucp18gm质粒为模板,以pel-yz-f和rhlyz-gc-r为引物扩增rhlyz基因片段,以palgc-pucp18gm质粒为模板,以rhlyz-gc-f和pel-gc-r为引物扩增algc基因片段,在通过overlappcr将rhlyz基因片段和algc基因片段进行融合,得到rhlyz-algc基因片段,通过无缝克隆拼接试剂盒与xhoi酶切的pk18-pel载体进行连接,转化e.colidh5α,得到自杀性质粒pk18-pel-rhlyz-algc。
(2)利用三亲结合的方法(受体菌株为pa111重组菌株,供体菌为pk18-pel-rhlyz-algc,助质粒为prk2013)将自杀性质粒转入到pa111重组菌株中,通过同源重组双交换,在敲除pel基因簇的同时将rhlyz-algc基因片段整合到染色体上,获得pa112重组菌株。
ppelup-f:agtaagcttcccttcgtttctggc(seqidno.35)
ppelup-r:attctcgagaccttcatgctcaatc(seqidno.36)
ppeldown-f:attctcgagcacggcgattcctttc(seqidno.37)
ppeldown-r:ataggatccacaccttggcgactctt(seqidno.38)
pel-yz-f:ctcagcgattgagcatgaaggtcagctggcacgacaggtttc(seqidno.39)
rhlyz-gc-r:gtcgggaaacctgtcgtgccagctgcttagacgctatggacgtg(seqidno.40)
rhlyz-gc-f:cacgtccatagcgtctaagcagctggcacgacaggtttcccgac(seqidno.41)
pel-gc-r:caagaaaggaatcgccgtgcatgtcagaagggcacgggca(seqidno.42)
3)pa113重组菌株的构建。构建过程如下:
(1)构建自杀性质粒pk18-alg-fadd4-lipc。以铜绿假单胞菌pao1为模板,分别以palgdup-f和palgdup-r、palgfdown-f和palgfdown-f扩增algd上游序列和algf下游序列;以实施例5中构建的pfadd4-pucp18gm质粒为模板,以palgd-fadd4-f和palgd-fadd4-r为引物扩增fadd4基因片段;以实施例4中构建的plipc-pucp18gm质粒为模板,以pfadd4-lipc-f和pfadd4-lipc-r为引物扩增lipc基因片段;同时将pk18gm载体用bamhi+hindiii进行双酶切并回收,将以上四个pcr产物进行纯化后与回收的载体通过无缝克隆拼接试剂盒进行连接,转化e.colidh5α,得到自杀性质粒pk18-alg-fadd4-lipc。
(2)利用三亲结合的方法(受体菌株为pa112重组菌株,供体菌为pk18-alg-fadd4-lipc,助质粒为prk2013)将自杀性质粒转入到pa112重组菌株中,通过同源重组双交换,在敲除alg基因簇的同时将fadd4和lipc基因整合到染色体上,获得pa113重组菌株。
palgdup-f:gaattcgagctcggtacccggggaaaagcccttgtggcgaatag(seqidno.43)
palgdup-r:caggtgggtttcaataacggtg(seqidno.44)
palgfdown-f:gaggtggtctgcctgtacgtcac(seqidno.45)
palgfdown-r:gtaaaacgacggccagtgccaagcttgcttcttcagttcttccaggtag(seqidno.46)
palgd-fadd4-f:caccgttattgaaacccacctgagctggcacgacaggtttc(seqidno.47)
palgd-fadd4-r:gaaacctgtcgtgccagctgatccagaggctcatgggaattc(seqidno.48)
pfadd4-lipc-f:gaattcccatgagcctctggatcagctggcacgacaggtttc(seqidno.49)
pfadd4-lipc-r:gtgacgtacaggcagaccacctctccaggtgccattagaccaaagac(seqidno.50)
4)pa114重组菌株的构建。构建过程如下:
(1)构建带有lac启动子的esta基因。以pucp18质粒为模板,以pk18esta-f1和pk18esta-r1为引物扩增lac启动子序列,以铜绿假单胞菌pao1为模板,以esta-f2和esta-r2为引物扩增esta基因,然后通过overlappcr,以pk18esta-f1和esta-r2为引物,将lac启动子序列和esta基因片段进行融合,并纯化,得到带有lac启动子的esta基因片段。
(2)构建自杀性质粒pk18-pha-esta。以铜绿假单胞菌pao1为模板,分别以phac1-f1和phac1-r1、phac2-f2和phac2-r2扩增phac1上游序列和phac2下游序列;同时将pk18gm载体用bamhi+hindiii进行双酶切并回收,将纯化后phac1上游序列、phac2下游序列以及带有lac启动子的esta基因片段与回收纯化的载体通过无缝克隆拼接试剂盒进行连接,转化e.colidh5α,得到自杀性质粒pk18-pha-esta。
(3)利用三亲结合的方法(受体菌株为pa113重组菌株,供体菌为pk18-pha-esta,助质粒为prk2013)将自杀性质粒转入到pa13重组菌株中,通过同源重组双交换,在敲除phac1-d-c2基因簇的同时将esta基因整合到染色体上,获得pa114重组菌株。
pk18esta-f1:acacgctgaacctgaatccagctggcacgacaggtttc(seqidno.51)
pk18esta-r1:gtggcttgagcgccattctgatcatggagctcgaattcgtaatcatggt(seqidno.52)
esta-f2:accatgattacgaattcgagctccatgatcagaatggcgctcaagccac(seqidno.53)
esta-r2:tgccaaggttgaagtcaggctgtacttcggtgttgaaactc(seqidno.54)
phac1-f1:gaattcgagctcggtacccgggagtccgatgcgatgcgtat(seqidno.55)
phac1-r1:gaaacctgtcgtgccagctggattcaggttcagcgtgt(seqidno.56)
phac2-f2:gagtttcaacaccgaagtacagcctgacttcaaccttggca(seqidno.57)
phac2-r2:gtaaaacgacggccagtgccaagctaacagcagggtcagggtgat(seqidno.58)
实施例8、pa111、pa112、pa113和pa114重组菌株与出发菌株pao1的鼠李糖脂合成及检测
实验重复三次,每次重复实验的步骤如下:
1)pa111、pa112、pa113和pa114重组菌株与出发菌株pao1的鼠李糖脂的合成
将pa111、pa112、pa113和pa114重组菌株与出发菌株pao1的单菌落分别接种于5mllb营养肉汤培养基(5g/l酵母粉、10g/l胰蛋白胨、10g/lnacl)中培养24h,次日以2%接种量转接至lb肉汤中培养16h,得到pa11、pa12、pa13和pa14重组菌株与出发菌株pao1的种子培养液。分别取800μl种子培养液,接种到含有15ml发酵培养基的三角瓶中[发酵培养基组成(perl):75gpalmoil;7.5gnano3;1.1g_kcl;1.1g_nacl;0.00028gfeso4·7h2o(1μm);3.4gkh2po4;4.4gk2hpo4;0.5gmgso4·7h2o;0.5gyeastextract;以及5ml微量元素溶液(perl):0.29gznso4·7h2o;0.24gcacl2·4h2o;0.25gcuso4·5h2o;and0.17gmnso4·h2o]。37℃,200rpm,培养96h。
2)pa111、pa112、pa113和pa114重组菌株与出发菌株pao1发酵液中鼠李糖脂的提取
分别取2mlpa111、pa112、pa113和pa114重组菌株与出发菌株pao1的发酵液,于12000rpm下离心5min,分别得到pa111、pa112、pa113和pa114重组菌株与出发菌株pao1发酵液上清,分别吸取1ml上清液,利用质量分数10%盐酸调ph至2.0;加入800μl乙酸乙酯,高速涡旋震荡20min进行萃取,10000g条件下离心5min吸出有机相,再次加入800μl乙酸乙酯萃取共三次,吸出三次萃取的有机相合并在一起于试管中;真空挥发有机相,待乙酸乙酯挥发完全后,得到膏状鼠李糖脂,向试管中加入5ml50mm的碳酸氢钠溶液,混合均匀,作为待测样品。(当鼠李糖脂产量较高时,适当调整加碳酸氢钠溶液量,并记录稀释倍数,以得到准确的吸光数值。)
3)pa111、pa112、pa113和pa114重组菌株与出发菌株pao1发酵液中鼠李糖脂的检测
(1)称重法检测鼠李糖脂
利用电子天平首先称量用于合并有机相的空试管重量(精确至0.1mg),待有机相挥发后再次称量有膏状鼠李糖脂试管的重量(精确至0.1mg),利用差减法计算出提取的粗鼠李糖脂的重量,随后该样品用于苔黑酚测定鼠李糖脂含量。
(2)苔黑酚法检测鼠李糖脂
以鼠李糖(国药集团化学试剂有限公司)为标准品,按照苔黑酚法做标准曲线定量分析鼠李糖脂,根据鼠李糖的含量计算pa111、pa112、pa113和pa114重组菌株与出发菌株pao1发酵液中的鼠李糖脂的量(rahmanpk,pasirayig,augerv,aliz(2009)productionofrhamnolipidbiosurfactantsbypseudomonasaeruginosads10-129inamicrofluidicbioreactor.biotechnolapplbiochem55(1):45-52),结果如图3所示。pao1菌株鼠李糖脂的合成量为22.6g/l,重组菌pa111、pa112、pa113和pa114的鼠李糖脂合成量分别为35.9、37.8、40.6和43.7g/l,尤其是pa114菌株,其鼠李糖脂产量较出发菌株pao1增加了93.4%(图3),纯度由65.7%提高到83.1%(图4),表明敲除鼠李糖脂前体合成旁路,同时过量表达有利于前体合成的基因使重组菌株鼠李糖脂产量明显增加。
(3)原始出发菌株pao1与重组菌株pa1114发酵液比较
出发菌株pao1在发酵过程中会产生更大更多的泡沫,发酵液也更粘稠,如图5a、5b所示。发酵液上清经乙酸乙酯抽提时,出发菌株pao1中间层很厚,上层有机相不易分离,而重组菌株pa1114中间层薄,上层有机相易分离(图5c)。吸取上层有机相并在真空干燥箱中使有机相挥发,得到黄褐色油脂状鼠李糖脂。由出发菌株提取的鼠李糖脂不透明,而经由重组菌株pa1114提取的鼠李糖脂澄澈透明(图5d),纯度较高,达到83.1%。
以上所述仅为本发明的较佳实施例,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!