一种玉米淀粉合成调控基因ZmMIKC2a及其应用的制作方法
本发明涉及的是基因工程的技术领域,尤其涉及的是一种玉米淀粉合成调控基因zmmikc2a及其应用。
背景技术:
玉米在人类的各类生产生活中占据着重要的位置,它不仅是主要的粮饲作物,更是各类生产中主要的淀粉来源。我国研究人员在提高玉米产量上做出了重要的贡献,但对改善玉米品质方面的研究较少淀粉作为植物种子胚乳的主要组成成分,其含量和品质直接影响着种子的品质。因此,研究淀粉合成的调控对于提高种子的品质具有重要意义。
淀粉根据结构不同分为直链淀粉和支链淀粉,淀粉中直链淀粉和支链淀粉的比例以及支链淀粉的分支链结构决定着玉米品质,同时直链淀粉和直链淀粉之间的比例还决定着淀粉的用途,高直链和低支链的淀粉可以用于食品、医药、环保、纺织等领域,而低直链和高支链的淀粉可用在食品行业。
目前,水稻、玉米、小麦淀粉代谢途径中控制四个关键酶(agpase、ss、sbe、dbe)的一些基因已经被克隆出来了,通过控制关键酶基因表达,能调控淀粉直链淀粉和支链淀粉含量的变化。在对高直链玉米酶活测定发现,ss和sbe酶活性低于普通玉米,而gbssi酶活性高于普通玉米。对控制高直链淀粉ae基因研究发现,与普通玉米比较,高直链玉米基因的第9个外显子缺失,cdna序列有碱基差异。在对小麦籽粒淀粉研究中发现,gbss在灌浆中前期与淀粉积累速率相关不显著,而灌浆后期呈显著相关;rna干涉水稻osbp-5基因能降低wx基因的表达水平,导致成熟种子中直链淀粉含量的降低;flo2(flouryendosperm2)基因能编码肽重复序列结构域,调控水稻籽粒的大小和淀粉质量;rsr1(ricestarchregulator1)基因在淀粉合成途径中起着负调控作用,该基因的缺失提高淀粉合成基因的表达,其突变体中直链淀粉增加,种子尺寸变大,产量提高;水稻的osbzip58基因,含有基本的亮氨酸拉链结构,能调控种子储藏蛋白的合成,同时还能绑定wx和sbei基因启动子,参与淀粉生物合成控制淀粉含量。尽管对于主效基因在淀粉合成途径中的功能也有大量报道,然而对于其分子调控机制仍不清楚。淀粉的含量是由多个主效基因控制的复杂生物学性状,那么这些主效基因是如何被调控表达?调控的分子机理又是怎么样的呢?这些问题都还不清楚。
技术实现要素:
本发明的目的在于克服现有技术的不足,提供了一种玉米淀粉合成调控基因zmmikc2a及其应用,以提供一种新的能够调控玉米合成相关基因表达水平,改变玉米直链淀粉与支链淀粉含量及其平衡度的调控基因。
本发明是通过以下技术方案实现的:
本发明提供了一种玉米淀粉合成调控基因zmmikc2a,所述基因具有如seqidno.1所示的核苷酸序列,全长810bp,共编码269个氨基酸。
本发明还提供了上述玉米淀粉合成调控基因zmmikc2a在调节玉米籽粒直链淀粉含量和玉米直链淀粉/总淀粉比例中的应用。
本发明还提供了上述玉米淀粉合成调控基因zmmikc2a的编码蛋白,所述编码蛋白具有如seqidno.2所示的氨基酸序列。
本发明还提供了一种植物过量表达载体,所述植物过量表达载体为多克隆位点区域依次连接有35s启动子、如权利要求1所述的zmend1a基因和终止子的pzz00005植物表达载体。
本发明还提供了一种遗传工程化的宿主细胞,所述宿主细胞含有上述过量表达载体,或其基因组中整合有外源的上述zmmikc2a基因的正向或反向序列。
本发明相比现有技术具有以下优点:本发明提供了一种玉米淀粉合成调控基因zmmikc2a及其应用,该基因能提高玉米直链淀粉的含量,改变籽粒中淀粉颗粒大小,调控淀粉合成相关基因表达水平;该基因的发现不仅为玉米淀粉合成调控机理提供新证据,也为创建玉米优质新品种奠定理论基础,为利用基因工程技术调节改善玉米直链和支链淀粉含量的平衡度提供了新的途径。
附图说明
图1为zmend1a基因不同组织表达模式分析结果柱状图;
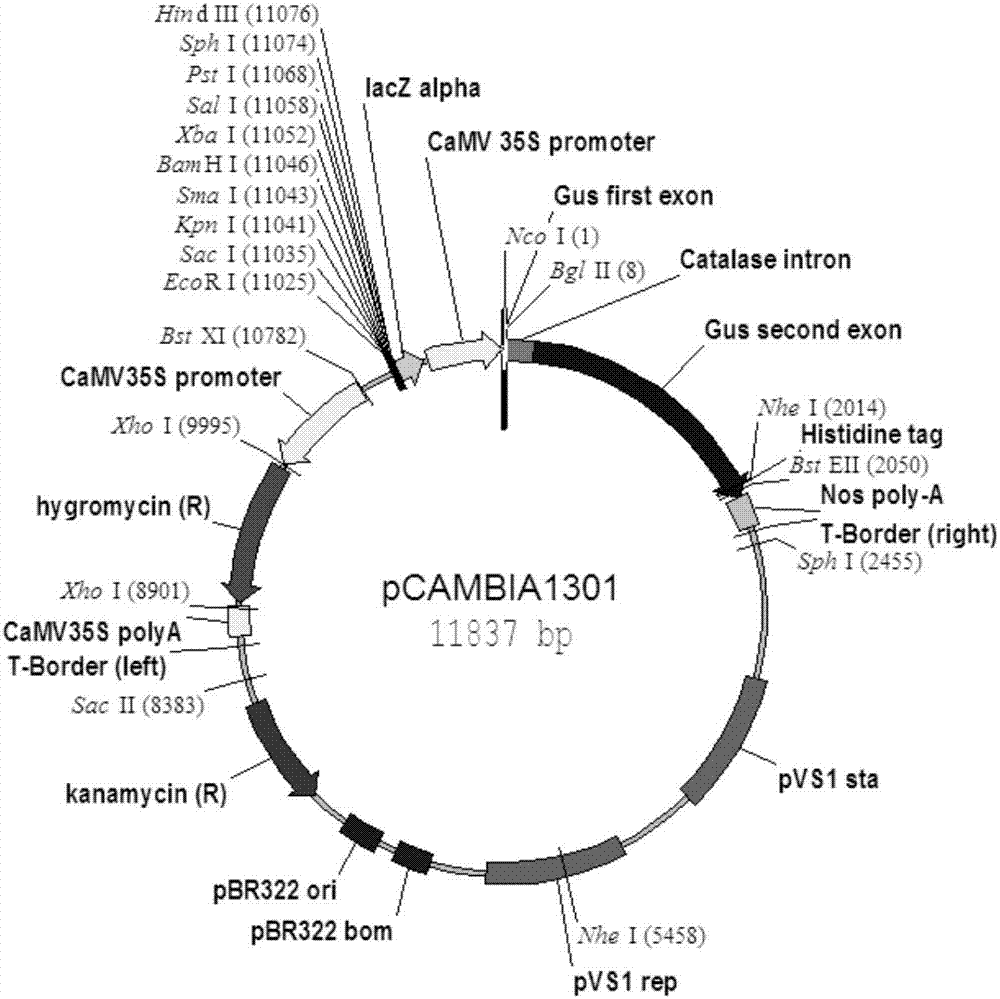
图2为基础载体图谱,其中,图2a为pcambia1301载体,图2b为pzz00005载体;
图3为zmmikc2a基因过表达植株和野生型植株籽粒大小和千粒重测定结果,其中图3a为籽粒大小对比图,图3b为千粒重测定结果柱状图;wt为野生型植株,l1-8为过表达植株;虚线为过表达植株千粒重均值;
图4为zmmikc2a基因过表达植株和野生型植株籽粒直链淀粉和总淀粉含量柱状图;其中图4a为直链淀粉含量柱状图,图4b为总淀粉含量柱状图;wt为野生型植株,l1、l2、l5、l8为过表达植株;虚线为过表达植株籽粒直链淀粉和总淀粉含量均值;
图5为zmmikc2a基因过表达植株和野生型植株籽粒扫描电镜图;其中wt为野生型植株,zmmikc2a为过表达植株;a为bar为100μm下的扫描电镜图,b为bar为30μm下的扫描电镜图。
图6为zmmikc2a基因过表达植株籽粒中淀粉合成相关基因表达分析结果图。
具体实施方式
下面对本发明的实施例作详细说明,本实施例在以本发明技术方案为前提下进行实施,给出了详细的实施方式和具体的操作过程,但本发明的保护范围不限于下述的实施例。
实施例1
1、材料
本实施例所用方法如无特别说明均为本领域的技术人员所知晓的常规方法,所用的试剂等材料,如无特别说明,均为市售购买产品。
2、方法
2.1zmmikc1a基因不同组织表达模式分析
分不同时期取玉米根(r)、茎(s)、叶(l)、雄穗(t)、花丝(c)、苞叶(st)、12d胚乳(en12)、12d胚(em12)、14d胚乳(en14)、14d胚(em14)、16d胚乳(en16)、16d胚(em16)、18d胚乳(en18)、18d胚(em18)、20d胚乳(en20)、20d胚(em20),每个样取完后,迅速放入液氮速冻,然后冻存于-70℃冰箱,用于后期rna时提取使用。rna的提取步骤参考e.z.n.a.tmmagsi植物rna提取试剂盒和使用核酸自动提取仪。荧光定量pcr引物见表1,以α-qtubulin(f:aggcttgtctcccaggtcatc;r:gttggtctggaactcgttcaca)作为内参基因,定量反应使用的是roche定量试剂盒,反应体系为25μl,各组分为染料混合液9.5μl,cdna模板为2μl,上下游引物各(10μmol/l)各0.5μl,最后补去离子水至25μl。pcr反应参数如下:95℃,10min;95℃,15s;60℃,1min,共40个循环。反应结束后,对产物进行加热,获得产物的溶解曲线。采用2–δδct[δct=ct目标基因–ct内参基因.δδct=δct处理后–δct对照]法进行数据处理。
实验测定三次生物学重复,至少三次实验操作重复,测定结果如图1所示,zmmikc1a基因在花丝、授粉后16天和授粉后18天表达量较高,特别是花丝中,表达量最高。
2.2zmmikc2a基因的克隆
选取玉米b73品种授粉后16天的籽粒胚乳,提取玉米胚乳rna并反转录成cdna,以玉米胚乳cdna为模板,根据玉米b73基因组数据库公布的基因序列,结合pcambia1301植物表达载体的多克隆位点设计引物,进行pcr扩增,获得pcr扩增产物。
引物序列为:
mikc2a-f-1:5′-cgcggatccatgggtaggggaaggattga-3′
mikc2a-r-1:5′-aactgcagttagaactgatgatgagggttattg-3′
pcr反应程序为:98℃,预变性10min;98℃,变性20s;60℃,退火20s;72℃,延伸2min,30个循环;72℃,复性10min;10℃保存。
将pcr扩增产物用质量比为2%的琼脂糖凝胶电泳检测,将与目的基因长度一致的电泳条带进行切胶回收,回收片段连接至peasy-t1载体(购于全式金生物技术有限公司,获得连接产物。将连接产物转化到大肠杆菌感受态trans5α细胞中,并提取质粒,以提取的质粒做模板,以mikc2a-f-1、mikc2a-r-1为引物进行pcr扩增验证,同时用bamhi和psti双酶切质粒进行检测,筛选阳性克隆子,将阳性克隆子送去华大生物公司进行测序,测序结果使用mega4.0软件比对,结果与预测一致。获得的重组质粒命名为t-mikc2a。
2.3zmmikc2a基因过量表达载体的构建
以pcambia1301(购于上海捷兰生物技术有限公司,载体图谱如图2a所示)为原始载体,在其多克隆位点的ecori和saci酶切位点之间连接一个35s启动子,并在sphi和hindiii酶切位点之间加上一段nos终止子,获得改造的载体p1。用bamhi和psti双酶切t-end1a得到小的目的片段,同时用bamhi和psti双酶切p1得到大的目的片段,将上述两片段使用t4连接酶连接,构建获得载体p1-zmmikc2a。
以p1-zmmikc2a为模板,以mikc2a-f-2和mikc2a-r-2为上下游引物进行pcr扩增,得到hindiii-35s+zmend1a+nos-pmei片段,分别用hindiii和pmei双酶切pzz00005基础载体(载体pzz00005购于中国种子集团有限公司,载体图谱如图2b所示)和hindiii-35s+zmend1a+nos-pmei片段,将上述两片段使用t4连接酶连接,构建获得载体p2-zmmikc2a。
引物序列为:
mikc2a-f-2:cccaagcttcgacactctcgtctactccaag
mikc2a-r-2:agctttgtttaaaccccgatctagtaacatagatgacac
2.4农杆菌介导的转zmmikc2a基因玉米的获得及鉴定
将p2-zmmikc2a转化农杆菌ag10中,获得重组农杆菌ag10/p2-zmmikc2a,具体参见文献:methodsinmolecularbiology,vol.343:agrobacteriumprotocols,2/e,volume1。
农杆菌介导对目的玉米的转化(玉米原位转化技术):
原位转化液的配制:1/4ms培养基中加2%(质量)蔗糖,0.05%(质量)silwet-77(表面活性剂),2mg/l6-ba,1ng/l2,4-d以及10mg/l乙酰丁香酮。每100ml培养基中加4ml农杆菌ag10/p2-zmmikc2a菌液,于28℃,250rpm培养24小时。
将玉米须剪去,用5ml的注射器将菌液从玉米的下部1/3处直接注入玉米棒中,注入的量以玉米须处有菌液渗出为宜。经过两个月的生长,收获玉米粒。
将玉米粒晒干,播种,待幼苗长到3-4叶期时用配制的0.01%(质量)除草剂basta和quickstixpat/bar蛋白检测试纸条(as013ls)检测表达载体p2-zmmikc2a上所含的bar基因蛋白,筛选阳性转基因植株,获得42株独立转化事件的玉米苗。
上述42株独立转化事件的玉米苗成熟后,以其叶片总dna为模板,使用quickstixpat/bar蛋白检测试纸条(as013ls)检测表达载体p2-zmmikc2a上所含的bar基因蛋白,同时使用zmmikc2a目的基因进行pcr检测(引物为mikc2a-f-1和mikc2a-r-1),鉴定转基因植株,鉴定时,以野生型目的玉米植株为阴性对照。
pcr反应程序为:94℃预变性10min;94℃变性30s,60℃退火20s,72℃延伸2min,30个循环;72℃总延伸10min。
结果,检测出18株阳性玉米植株,即为t0代转基因玉米植株,命名为l1-18。t0代转基因玉米植株成熟后,收获t1代转基因玉米种子,t1代转基因玉米种子自交繁殖得到t2代转基因玉米种子。依次类推,获得zmmikc2a基因过量表达的转基因玉米株系。
2.5zmmikc2a基因过表达植株的籽粒质量检测分析
2.5.1籽粒大小和百粒重测定
取8株独立转化事件转基因株l1-8的t3代籽粒进行大小和百粒重测定,并以野生型玉米wt籽粒做对照。
结果如图3所示,其中,图4a可看出,转基因玉米籽粒大小变化不明显,图4b可看出,转基因玉米籽粒千粒重除l1和l5稍小外,其它几个株系和野生型无明显差别。
2.5.2直链淀粉和总淀粉含量的测定
取4株独立转化事件转基因株l1、l2、l5、l8的t3代籽粒进行直链淀粉和总淀粉测定,并以野生型玉米籽粒做对照,每次实验测定至少重复三次,测定方法如下:
直链淀粉测定时,首先对籽粒中淀粉进行提纯,将烘干去硬壳和胚的玉米种子在室温下用0.4%(质量分数)的naoh溶液浸泡48h,料液(样本比0.4%的naoh溶液)比为1:3,洗涤后再用0.4%的naoh溶液浸泡24h,反复水洗至种子表面不再黏滑为止,沥干水分,用胶体磨粉碎,淀粉乳过筛(200目),离心(3000r/min,20min),取下层白色沉淀,即为淀粉,将淀粉反复漂洗离心,40℃鼓风干燥,粉碎,过筛(200目),即得淀粉纯品,用于直链淀粉的测定。直链淀粉的测定采用爱尔兰megazyme公司生产的直链淀粉/支链淀粉分析试剂盒(货号:k-amyl),测定方法可参见试剂盒说明书,在此不做详细描述。
总淀粉测定时,先将烘干的种子去硬壳和胚,再使用组织研磨仪将其研磨成粉末,40℃过夜烘干,过0.5mm筛(35目)。总淀粉测定采用爱尔兰megazyme公司生产的总淀粉测定试剂盒(货号:k-tsta),具体为:将研磨后样品(准确称量~100mg)加入到试管中(16×120mm),确保全部样品位于试管底部;加入0.2ml乙醇溶液(80%v/v),用漩涡混合器混合;放入搅拌架子,加入2ml的2mol/l的koh至每个试管中,冰水混合浴中搅拌20min左右,重悬粉末和溶解抗性淀粉(注意:不要使用涡旋仪混匀,会使淀粉乳化,确保加入koh时,试管处于剧烈震荡状态。这是为了避免淀粉形成团块难以溶解);当试管在磁力搅拌器上搅拌时,加入8ml的1.2mol/l醋酸钠缓冲液(ph3.8)至每个试管。立即加入0.1ml耐热α-淀粉酶(试剂盒配置溶液)和0.1ml淀粉葡糖苷酶(试剂盒配置溶液),充分混合,在50℃下孵育;孵育30min,间断混匀;样品淀粉含量>10%:将全部的溶液转移到100ml的容量瓶中,用洗瓶冲洗试管,并也倒入容量瓶中,然后用蒸馏水调整溶液体积至100ml,充分混匀。取部分溶液离心(1800rpm,10min);转移两份稀释的样品溶液(0.1ml)到圆底试管中(16×100mm);加入3.0mlgopod溶液(试剂盒配置溶液)到每一个试管中(包括葡萄糖对照和试剂空白对照),然后在50℃下孵育20min;葡萄糖对照包括0.1ml葡萄糖标准溶液(1mg/ml)+3.0mlgopod溶液。试剂空白对照:0.1ml蒸馏水+3.0mlgopod溶液;相对于试剂空白在510nm下测定每一个样品和葡萄糖质控的吸光度。计算公式如下:
δa=相对于试剂空白所读取的吸光度;fv=总体积(例如100ml或10ml);0.1=样品分析体积;1/1000=转化从μg至mg;100/w=淀粉作为干粉重量的比例;w=干粉重量;162/180=d葡萄糖转化为脱氢葡萄糖;淀粉%(占干物质比重)=淀粉%×100/100-水分含量(%)。
测定结果如图4所示,图中可以看出,zmmikc2a转基因玉米籽粒中的直链淀粉含量提高显著,幅度在3.93%-11.42%(图4a),总淀粉含量几乎不变(图4b),直链淀粉/总淀粉比例显著上调。该结果说明了zmmikc2a基因能直接或间接影响淀粉合成过程中直链淀粉相关基因的表达,对直链淀粉表达起正调控作用。
2.5.3玉米籽粒淀粉颗粒扫描电镜观察
扫描电镜具体实验操作方法如下:
①取晒干的种子,去壳;使用锋利的切割刀,将种子从中间纵切成两半,分别标号装到离心管中;
②向离心管中加入质量比2.5%的戊二醛溶液,4℃过夜固定,倒掉固定液后,用0.1mol/l的ph7.0的磷酸缓冲液漂洗种子3次,每次15min;再用质量比1%的锇酸溶液固定,常温下,1-2h,倒掉固定液后,用0.1mol/l的ph7.0磷酸缓冲液漂洗种子3次;
③从离心管中吸去磷酸缓冲液,加入梯度浓度的乙醇溶液分别对种子进行脱水处理。乙醇梯度浓度(质量比)分别为:30%,50%,70%,80%和90%,每种浓度处理20min,再用纯乙醇洗脱2次,每次20min;
④吸去离心管中的纯乙醇,用乙醇和醋酸异戊酯混合液(v:v=1:1)处理种子30min(间隔摇动溶液),再用纯醋酸异戊酯溶液处理种子1-2h;
⑤使用临界干燥法,将种子从醋酸异戊酯溶液中挑到样品笼中,用滤纸吸去样品笼外围的溶液后,移入到仪器的样品杯中,盖盖,拧紧(注:在装样前,先打开仪器的电源,将温度设定为0℃预处理10-15min);
⑥打开co2排气阀和仪器的进气阀,向样品杯中注入液体co2,当co2的量到样品杯的50%时,关闭进气阀,静止15-20min;
⑦再打开仪器排气流量计阀门和进气阀,10min后关闭排气阀;当充入的co2到样品杯的80%左右时,关闭进液阀;
⑧加温置换:将仪器温度调至20℃,放置15-20min后,杯中的co2会逐渐气化,当杯中的气压升到约为7000pa时,醋酸异戊酯会与co2充分置换;
⑨气化:将温度调至35-40℃时,杯内的co2达到临界点;当压力达到7134pa时,5min后,打开流量计的排气阀,以1.0-1.51/min的速度排气,45-60min后结束排气,样品杯的压力降为0,当温度降至室温后,即可取出样品;
⑩镀膜:将种子非观察面,用导电胶粘贴到样品台上,胶风干后,进行镀膜,本实验采用离子溅射镀膜法:装置由真空泵和真空罩两部分组成,真空罩内有阴阳极,阴极面装黄金靶,种子装在阳极面的样品座上。当真空罩中的真空度抽到1-10pa时,在阴阳极之间加上2000v的直流电压,此时会产生弧光放电电场(在电场的作用下,真空罩里的残余气体分子会被电离为正离子和电子,正离子被阳极吸引轰击黄金靶,激发出黄金颗粒和电子,并被阳极吸引附着在种子表面形成导电膜);置于扫描电镜下观察,拍照。
扫描电镜结果如图5所示,图中可发现,转基因玉米淀粉颗粒小于野生型材料,说明zmmikc2a基因过量表达减小了玉米籽粒中淀粉颗粒尺寸。
2.5.4转基因玉米籽粒中相关淀粉合成基因的表达变化分析
分别取野生型玉米和转基因玉米t3代植株授粉后5d、10d、15d、20d和25d的种子,样品取完后立即放入液氮速冻,冻存于-80℃冰箱中,备用。利用e.z.n.a.tmmagsi植物总rna提取试剂盒提取样品rna,并反转录成cdna,以cdna为模板,以a-tubulin为内参基因,利用roche荧光定量试剂盒,选择11种已知的玉米淀粉合成相关基因进行实时荧光定量pcr检测,具体为:
荧光定量pcr扩增使用的引物:
①内参基因引物:
a-tubulin-f:5’-gggattgccgatcgtatgag-3’
a-tubulin-r:5’-agccaccgatccagacact-3’
②荧光定量pcr扩增玉米淀粉合成相关基因的引物:
荧光定量pcr反应体系为:
荧光定量pcr反应程序为:95℃预变性10min,95℃变性15s,60℃退火1min,共40个循环。反应结束后,对产物进行加热,获得产物的溶解曲线。采用2–δδct[δct=ct目标基因–ct内参基因.δδct=δct处理后–δct对照]法进行数据处理。
以野生型材料授粉后5d的表达量作为对照,每次基因6次生物学重复和3次实验操作,结果如图6所示,图中可以看出,zmagpl1在整个时期表达水平都高于对照组材料;zmagps1a-1、zmbeiib、zmisa1、zmssiia、zmssiiia和zmgbssia在10-20d时间内表达水平高于对照组材料;zmbeiia、zmgbssiia和zmpul在20d时表达水平高于对照组材料,其它时期不变;而zmphol基因除15d表达水平高于对照组材料外,其它时期都低于对照组材料。上述结果进一步说明了,zmmikc1a基因的转入对淀粉合成相关基因产生了很大的影响,大部分基因表达水平提高,从而使籽粒中淀粉含量和淀粉颗粒发生改变。
以上为本发明一种详细的实施方式和具体的操作过程,是以本发明技术方案为前提下进行实施,但本发明的保护范围不限于上述的实施例。
sequencelisting
<110>安徽省农业科学院烟草研究所
<120>一种玉米淀粉合成调控基因zmmikc2a及其应用
<130>/
<160>2
<170>patentinversion3.3
<210>1
<211>810
<212>dna
<213>zeamays
<400>1
atgggtaggggaaggattgagatcaagaggatcgagaacaacacgagccggcaggtcacc60
ttctgcaagcgccgcaatgggctcctcaagaaggcgtatgagctctccgtcctctgcgac120
gctgaggtggccctcatcgtcttctctagccgtggtcgcctctacgagtatgccaacaac180
agtgtcaaggctactattgagaggtacaagaaggcacacaccgttggctcttcctctggg240
ccaccgctcctagagcacaatgcccagcaattctaccagcaagaatcagcaaaactgcgc300
aaccagatccagatgctgcaaaacactaacaggcacttggttggtgattccgtgggaaac360
ctgtcactcaaggagctgaagcagctggagagccgccttgagaaaggcatctctaagatc420
agggccaggaagagtgagctgctggctgcggagatcagttacatggccaaaagggagact480
gagcttcagaatgaccacatgaccctcaggaccaagattgaggagggagagcaacagctg540
cagcaggtgaccgtggcacggtcagttgcagcagcagcagcagctgccaccaacttggag600
ctgaacccattcttagagatggataccaaatgcttcttcactggcggccccttcgcaacg660
ctggacatgaagtgctttctccccggcagcttgcagcagatgctggaggcacagcagcgc720
cagatgctcgccaccgagctgaacctcggctaccaactggcgccgcctggttctgacgct780
gccgacaataaccctcatcatcagttctaa810
<210>2
<211>269
<212>prt
<213>zeamays
<400>2
metglyargglyargilegluilelysargilegluasnasnthrser
151015
argglnvalthrphecyslysargargasnglyleuleulyslysala
202530
tyrgluleuservalleucysaspalagluvalalaleuilevalphe
354045
serserargglyargleutyrglutyralaasnasnservallysala
505560
thrilegluargtyrlyslysalahisthrvalglysersersergly
65707580
proproleuleugluhisasnalaglnglnphetyrglnglngluser
859095
alalysleuargasnglnileglnmetleuglnasnthrasnarghis
100105110
leuvalglyaspservalglyasnleuserleulysgluleulysgln
115120125
leugluserargleuglulysglyileserlysileargalaarglys
130135140
sergluleuleualaalagluilesertyrmetalalysarggluthr
145150155160
gluleuglnasnasphismetthrleuargthrlysilegluglugly
165170175
gluglnglnleuglnglnvalthrvalalaargservalalaalaala
180185190
alaalaalaalathrasnleugluleuasnpropheleuglumetasp
195200205
thrlyscysphephethrglyglyprophealathrleuaspmetlys
210215220
cyspheleuproglyserleuglnglnmetleuglualaglnglnarg
225230235240
glnmetleualathrgluleuasnleuglytyrglnleualapropro
245250255
glyseraspalaalaaspasnasnprohishisglnphe
260265
- 还没有人留言评论。精彩留言会获得点赞!
- 拟南芥SSCD1基因突变在调控植物体内茉莉酸合成中的应用的制作方法与工艺
- 一种基因调控网络的优化控制方法与流程
- 拟南芥WRKY家族转录因子AtWRKY44基因,蛋白编码序列及其应用的制作方法与工艺
- 针对免疫调控能力相关基因个体化配方食品及其制备方法与流程
- REVOLUTA基因在调控豆科植物小叶片数目和叶茎比中的应用的制作方法与工艺
- 玉米锌铁调控转运体ZmZIP3基因及其应用的制作方法与工艺
- 拟南芥基因SPOC1在调控植株开花期中的应用的制作方法与工艺
- 调控石榴花胚珠发育的PgAG基因及其用途的制作方法与工艺
- 光效调控基因HPE1及其应用的制作方法与工艺
- 菘蓝IiAP2/ERF049基因在调控木脂素类化合物合成中的应用的制作方法与工艺